Типы побегов. Органы растения: побег
Зародыша либо из пазушной или придаточной (адвентивной) почки. Таким образом, почка является зачаточным побегом. При прорастании семени из зародышевой почечки формируется первый побег растения - его главный побег , или побег первого порядка .
Из главного побега формируются боковые побеги , или побеги второго порядка , а при повторении ветвления - третьего порядка и т. д.
Придаточные побеги формируются из придаточных почек.
Так формируется система побегов, представленная главным побегом и боковыми побегами второго и последующего порядков. Система побегов увеличивает общую площадь соприкосновения растения с воздушной средой.
В зависимости от выполняемой функции различают побеги вегетативные, вегетативно-генеративные и генеративные. Вегетативные (невидоизменённые) побеги, состоящие из стебля, листьев и почек и вегетативно-генеративные (частично видоизменённые), состоящие дополнительно из цветка или соцветия, выполняют функции воздушного питания и обеспечивают синтез органических и неорганических веществ. В генеративных (полностью видоизменённых) побегах фотосинтез чаще всего не происходит, зато там образуются спорангии, задача которых сводится к обеспечению размножения растения (к таким побегам относится и цветок).
Побег, на котором образуются цветки, называется цветоносным побегом , или цветоносом (иногда термин «цветонос» понимают в более узком смысле - как участок стебля, на котором находятся цветки).
Основные органы побега
Вегетативный невидоизменённый побег - единый орган растения, состоящий из стебля, листьев и почек, формирующийся из общего массива меристемы (конуса нарастания побега) и обладающие единой проводящей системой. Стебли и листья, являющиеся основными структурными элементами побега - часто рассматриваются как его составные органы, то есть органы второго порядка. Кроме того, обязательная принадлежность побега - почки. Главная внешняя черта, отличающая побег от корня - наличие листьев.
Моноподиальное ветвление
Моноподиальное ветвление является следующим этапом эволюции ветвления побегов. У растений с моноподиальным типом строения побега верхушечная почка сохраняется на протяжении всей жизни побега. Моноподиальный тип ветвление часто встречается среди голосеменных растений, также встречается у многих покрытосеменных (например, у многих видов пальм, а также растений из семейства Орхидные - гастрохилусов, фаленопсисов и других). Некоторые из них имеют единственный вегетативный побег (например, Фаленопсис приятный).
Моноподиальные растения - термин, наиболее часто используемый в описании растений тропической и субтропической флоры, а также в научно-популярной литературе по комнатному и оранжерейному цветоводству.
Моноподиальные растения могут существенно различаться внешне. Среди них есть розеточные, с удлиненным побегом, кустовидные.
Симподиальное ветвление
У растений с симподиальным типом строения побега верхушечная почка закончив развитие отмирает или дает начало генеративному побегу . После цветения этот побег больше не растет, а у его основания начинает развиваться новый. Структура побега у растений с симподиальным типом ветвления сложнее, чем у растений с; симподиальное ветвление является эволюционно более продвинутым типом ветвления. Слово «симпоидальный» образовано от греч. sym («вместе» или «много») и pod («нога»).
Симподиальное ветвление характерно для многих покрытосеменных растений: например, для лип, ив и многих орхидных.
У орхидных, помимо верхушечных, у части симподиальных орхидей образуются и боковые соцветия, развиваясь из почек, расположенных у основания побега (Пафиния гребенчатая). Часть побега, прижатая к субстрату, назвается ризомой. Располагается, как правило, горизонтально и не имеет настоящих листьев, только чешуевидные. Редуцированной, почти неразличимой ризома бывает у многих масдеваллий, дендробиумов и онцидиумов; хорошо различимой и утолщенной - у каттлей и лелий, удлиненной - у бульбофиллюмов и целогин, достигая 10 и более сантиметров. Вертикальная часть побега часто бывает утолщенной, образующей так называемый туберидий, или псевдобульбу. Псевдобульбы могут быть различной формы - от почти сферической до цилиндрической, конусовидной, булавовидной и удлиненной, напоминающей стебли тростника. Псевдобульбы являются запасающими органами.
Симподиальные растения - термин, наиболее часто используемый в описании растений тропической и субтропической флоры, а так же в научно-популярной литературе по комнатнуму и оранжерейному цветоводству.
|
Bulbophyllum grandiflorum |
Cirrhopetalum macraei |
Oncidium dasystyle . Curtis"s botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 г. |
Dendrobium senile . Curtis"s botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 г. |
Эволюция типов ветвления
Видоизменения побегов (метаморфоз)
Побег - самый изменчивый по внешнему облику орган растения. Это связано не только с общей мультифункциональностью вегетативных органов , возникшей в процессе эволюции, но и с изменениями, происходящими в процессе онтогенеза растений, обусловленными адаптацией к разнообразию условий окружающей среды, а у культурных растений - под воздействием человека.
Формируется корневище или изначально как подземный орган (купена, вороний глаз, ландыш, черника), или сначала как надземный ассимилирующий побег, который затем погружается в почву с помощью втягивающих корней (земляника, медуница, манжетка). Корневища могут нарастать и ветвиться моноподиально (манжетка, вороний глаз) или симподиально (купена, медуница). В зависимости от длины междоузлий и интенсивности нарастания, различают длинные и короткие корневища и, соответственно, длиннокорневищные и короткокорневищные растения.
При ветвлении корневищ образуется куртина надземных побегов, связанных участками системы корневищ. Если связующие части разрушаются, побеги обособляются, и происходит вегетативное размножение. Совокупность новых особей, образовавшихся вегетативным путем, называют клоном . Корневища характерны преимущественно для травянистых многолетников, но встречаются и у кустарников (бересклет) и кустарничков (брусника, черника).
Близки к корневищам подземные столоны – недолговечные тонкие подземные побеги, несущие недоразвитые чешуевидные листья. Столоны служат для вегетативного размножения, расселения и захвата территории. Запасные питательные вещества в них не откладываются.
У некоторых растений (картофель, земляная груша) к концу лета из верхушечных почек столонов образуются клубни (рис. 4.24 ). Клубень имеет шаровидную или овальную форму, стебель сильно утолщен, в нем откладываются запасные питательные вещества, листья редуцируются, в их пазухах формируются почки. Столоны отмирают и разрушаются, клубни перезимовывают, и на следующий год дают начало новым надземным побегам.
Клубни не всегда развиваются на столонах. У некоторых многолетних растений клубневидно разрастается и утолщается основание главного побега (цикламен, капуста кольраби) (рис. 4.24 ). Функции клубня – запас питательных веществ, переживание неблагоприятного периода года, вегетативное возобновление и размножение.
У многолетних трав и полукустарничков с хорошо развитым стержневым корнем, сохраняющимся в течение всей жизни, образуется своеобразный орган побегового происхождения, называемый каудексом . Вместе с корнем он служит местом отложения запасных веществ и несет множество почек возобновления, часть из которых могут быть спящими. Каудекс обычно бывает подземным и образуется из коротких оснований побегов, погружающихся в почву. От коротких корневищ каудекс отличается способом отмирания. Корневища, нарастая верхушкой, постепенно отмирают и разрушаются на более старом конце; главный корень не сохраняется. Каудекс разрастается в ширину, с нижнего конца он постепенно переходит в долгоживущий утолщающийся корень. Отмирание и разрушение каудекса и корня идет от центра к периферии. В центре образуется полость, а затем он может разделиться продольно на отдельные участки – партикулы . Процесс разделения особи стержнекорневого растения с каудексом на части называют партикуляцией . Каудексовых растений много среди бобовых (люпин, люцерны), зонтичных (бедренец, ферула), сложноцветных (одуванчик, полыни).
Луковица – это, как правило, подземный побег с очень коротким уплощенным стеблем – донцем и чешуевидными мясистыми сочными листьями, запасающими воду и растворимые питательные вещества, главным образом сахара. Из верхушечной и пазушных почек луковиц вырастают надземные побеги, на донце образуются придаточные корни (рис. 4.24 ). Таким образом, луковица – типичный орган вегетативного возобновления и размножения. Луковицы наиболее характерны для растений из семейств лилейных (лилии, тюльпаны), луковых (луки) и амариллисовых (нарциссы, гиацинты).
По структуре луковицы очень разнообразны. В одних случаях запасающие чешуи луковицы представляют собой только видоизмененные листья , не имеющие зеленых пластинок (лилия саранка); в других – это подземные влагалища зеленых ассимилирующих листьев, которые утолщаются и остаются в составе луковицы после отмирания пластинок (лук репчатый). Нарастание оси луковицы может быть моноподиальным (подснежник) или симподиальным (гиацинт). Наружные чешуи луковицы расходуют запас питательных веществ, подсыхают и играют защитную роль. Число луковичных чешуй варьирует от одной (чеснок) до нескольких сотен (лилии).
В качестве органа возобновления и запаса луковица приспособлена главным образом к климатам средиземноморского типа – с достаточно мягкой влажной зимой и с очень жарким засушливым летом. Она служит не столько для благополучной перезимовки, сколько для переживания жесткой летней засухи. Запасание воды в тканях луковичных чешуй происходит за счет образования слизей, способных удерживать большое количество воды.
Клубнелуковица внешне напоминает луковицу, но ее чешуевидные листья не являются запасающими; они сухие и пленчатые, а запасные вещества откладываются в утолщенной стеблевой части (шафран, гладиолус).

Рис. 4.24. Подземные метаморфозы побега : 1, 2, 3, 4 – последовательность развития и строение клубня картофеля; 5 – клубень цикламена; 6 – клубень кольраби; 7 – луковички лилии тигровой; 8 – луковица лука репчатого; 9 – луковица лилии; 10 – участок длинного корневища пырея ползучего.
Видоизменяться могут не только подземные, но и надземные побеги растений (рис. 4.25 ). Довольно часто встречаются надземные столоны . Это плагиотропные недолговечные побеги, функция которых вегетативное размножение, расселение и захват территории. Если столоны несут зеленые листья и участвуют в процессе фотосинтеза, их называют плетями (костяника, живучка ползучая). У земляники столоны лишены развитых зеленых листьев, стебли у них тонкие и хрупкие, с очень длинными междоузлиями. Такие более высокоспециализированные для функции вегетативного размножения столоны называют усами .
Сочными, мясистыми, приспособленными для накопления воды могут быть не только луковицы, но и надземные побеги, обычно у растений, живущих в условиях недостатка влаги. Водозапасающими органами могут быть листья или стебли, иногда даже почки. Такие сочные растения носят название суккулентов . Листовые суккуленты запасают воду в тканях листьев (алоэ, агавы, толстянки, родиола, или золотой корень). Стеблевые суккуленты характерны для американского семейства кактусовых и африканских молочайных. Сочный стебель выполняет водозапасающую и ассимилирующую функцию; листья редуцированы или превращены в колючки (рис. 4.25, 1). У большинства кактусов стебли колонновидные или шаровидные, листьев на них совсем не образуется, но узлы хорошо заметны по расположению пазушных побегов – ареол , имеющих вид бородавок или удлиненных выростов с колючками или пучками волосков. Превращение листьев в колючки уменьшает испаряющую поверхность растения и защищает от поедания животными. Примером метаморфоза почки в суккулентный орган – кочан служит культурная капуста.
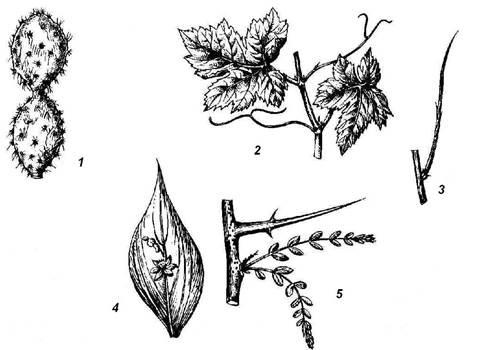
Рис. 4.25. Надземные метаморфозы побега: 1 – стеблевой суккулент (кактус); 2 – усики винограда; 3 – безлистный фотосинтезирующий побег дрока; 4 – филлокладий иглицы; 5 – колючка гледичии.
Колючки кактусов имеют листовое происхождение. Листовые колючки нередко встречаются и у несуккулентных растений (барбарис) (рис. 4.26, 1). У многих растений колючки имеют не листовое, а стеблевое происхождение. У дикой яблони, дикой груши, жостера слабительного в колючки метаморфизируются укороченные побеги, имеющие ограниченный рост и оканчивающиеся острием. Вид жесткой одревесневшей колючки они приобретают после опадения листьев. У боярышника (рис . 4.26, 3) колючки, образующиеся в пазухах листьев, совершенно безлистны с самого начала. У гледичии (рис. 4.25, 5 ) мощные разветвленные колючки образуются на стволах из спящих почек. Образование колючек любого происхождения, как правило, есть результат недостатка влаги. При выращивании многих колючих растений в искусственной влажной атмосфере они утрачивают колючки: вместо них вырастают нормальные листья (верблюжья колючка) или облиственные побеги (английский дрок).
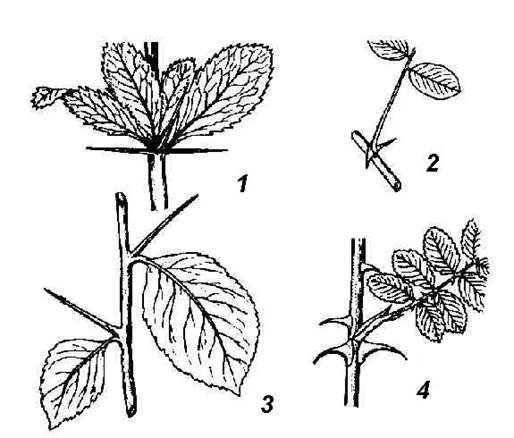
Рис. 4.26. Колючки различного происхождения : 1 – листовые колючки барбариса; 2 – колючки белой акации, видоизменение прилистников; 3 – колючки боярышника побегового происхождения; 4 – шипы – эмергенцы шиповника.
Побеги ряда растений несут шипы . Шипы отличаются от колючек меньшими размерами, это выросты – эмергенцы – покровной ткани и тканей коры стебля (шиповник, крыжовник) (рис. 4.26, 4 ).
Приспособление к недостатку влаги очень часто выражается в ранней утрате, метаморфозе или редукции листьев, теряющих основную функцию фотосинтеза. Это компенсируется тем, что роль ассимилирующего органа берет на себя стебель. Иногда такой ассимилирующий стебель безлистного побега остается внешне неизмененным (испанский дрок, верблюжья колючка) (рис. 4.25, 3). Дальнейшим шагом в указанной смене функций является формирование таких органов, как филлокладии и кладодии . Это уплощенные листоподобные стебли или целые побеги. На побегах иглицы (рис. 4.25, 4 ), в пазухах чешуевидных листьев, развиваются плоские листовидные филлокладии, имеющие, подобно листу, ограниченный рост. На филлокладиях образуются чешуевидные листья и соцветия, чего никогда не бывает на нормальных листьях, значит, филлокладий соответствует целому пазушному побегу. Мелкие, игольчатые филлокладии образуются у спаржи в пазухах чешуевидных листьев основного скелетного побега. Кладодии – это уплощенные стебли, сохраняющие, в отличие от филлокладиев, способность к длительному росту.
Для некоторых растений характерно видоизменение листьев или их частей, а иногда целых побегов в усики , которые закручиваются вокруг опоры, помогая тонкому и слабому стеблю сохранять вертикальное положение. У многих бобовых в усики превращается верхняя часть перистосложного листа (горох, горошки, чина). В других случаях в усики превращаются прилистники (сассапариль). Очень характерные усики листового происхождения формируются у тыквенных, причем можно видеть все переходы от нормальных листьев к полностью метаморфизированным. Усики побегового происхождения можно наблюдать у винограда (рис. 4.25, 2), пассифлоры и ряда других растений.
Надземную часть растений называют термином "побег". Строение его обусловлено выполняемыми функциями. Безусловно, каждый орган незаменим и определяет возможность существования биологического вида. Функции питания, процессы роста, возможность адаптации - вот только некоторые из важнейших функций видимой части растительных организмов.
Биология: строение побега
В морфологии различают осевую и боковую части данного органа: стебель и лист. Строение поражает своим многообразием: от микроскопической водной ряски до гигантской лесной секвойи. Оно обусловлено различным строением составных частей надземной части, связанных с особенностями среды обитания и климатическими условиями. Здесь также находятся укороченные зачаточные побеги - почки.
Место прикрепления листа к осевой части называется узлом, а угол, который образуется между ними, - пазухой. Здесь располагаются специализированные почки, образующие листья или цветки. Расстояние между двумя местами крепления листьев называется междоузлием.
Стебель
Строение побега первоначально зависит от направления роста и расположения в пространстве стебля. В зависимости от этих признаков различают прямостоячие, стелющиеся, ползучие, вьющиеся и цепляющиеся виды. Разнообразны стебли и характером поверхности. Она может быть голой или с выростами, гладкой или шероховатой. Если разрезать стебель поперек, то можно определить форму: округлую, ребристую, с определенным количеством граней или сплюснутую.
![]()
Усы земляники также являются ее побегом, имеющим укороченные междоузлия.
В зависимости от жизненной формы различают травянистые и деревянистые стебли. Первые не имеют камбия - боковой В первые годы жизни внешне на них похожи новые побеги деревьев и кустарников. Они зеленого цвета и способны к фотосинтезу. С течением времени они одревесневают, становясь более прочными. Они способны удерживать крупные плоды и противостоять сильным порывам ветра.
Виды стеблей
В зависимости от особенностей цикла развития растения могут быть одно-, дву- и многолетние. Например, астры отцветают осенью, после чего полностью отмирают. По-другому развивается морковь и свекла. На первом году жизни они формируют корнеплоды, которые представляют собой органы, запасающие питательные вещества. Осенью их стебель погибает. Но растение существует в виде видоизмененного стебля. С наступлением благоприятных условий побег снова вырастает. При этом на второй год жизни на нем в результате цветения образуются семена, с помощью которого растение размножается.

Цикл жизни многолетних растений рассмотрим, изучив особенности строения побегов хвойных растений . Это кустарники или деревья, имеющие единственный мощный стебель - ствол. Его развитие начинается с прорастания семени. В результате его роста образуется проросток, а после - взрослое растение. Заканчивается жизненный цикл многолетних растений гибелью. Хвойные по праву считаются настоящими долгожителями. Так, сосна живет около 400 лет, ель - до 500, а можжевельник - целых 1000!
Лист
Боковая часть побега не менее функциональна и разнообразна. Она обеспечивает воздушное питание, транспирацию - испарение воды с поверхности, вегетативное размножение. Побег, строение которого обусловловлено выполняемыми функциями, характеризуется разнообразными листьями.
Иголки кактуса нужны, чтобы сократить количество испаряемой влаги. А пальчатые широкие у конского каштана, наоборот, увеличивают ее количество.
Листья с одной пластинкой называются простыми, а с несколькими, расположенными на одном черешке, - сложными. Рассматривая их, можно заметить определенный рисунок. Его создают жилки. Это сосудисто-волокнистые пучки По характеру жилкования различают листья с сетчатым (клен, яблоня), параллельным (кукуруза, рожь) и дуговым (подорожник, ландыш) жилкованием.
Листорасположение
Побег, строение которого зависит и от количества получаемой солнечной энергии, характеризуется различным расположением листьев на стебле. В случае их расположения по спирали образуется очередное, а если по кругу - супротивное, или мутовчатое.
В природе не существует растений, которые не обновляют листву. Ее сбрасывает и сосна, и ель. Поскольку опадают не все листья сразу, этого никто не замечает.

Видоизменения побега
При возникновении необходимости выполнения дополнительных функций побеги и их составные части видоизменяются. Листья могут превращаться в колючки или чешуйки. У растительных хищников они способны улавливать и переваривать мелких насекомых.

Клубни топинамбура, который также называют земляной грушей, также образуют видоизменение побега - клубень. На утолщенном мясистом стебле находятся рубцевидные почки, из которых вырастают молодые побеги.
Подземными стеблями с удлиненными междоузлиями являются корневища. Они похожи на плети, имеют хорошо развитые механические и проводящие ткани. Из почек, расположенных на корневище, образуются листья. Те, кто плохо знаком с особенностями строения корневищ, считают, что если избавиться от листьев, все растение погибнет. Но это мнение ошибочно, ведь основная часть растения надежно защищена и находится под землей.
Взаимосвязь строения и функций
Строение побега зависит от выполняемых функций. Это можно доказать, рассмотрев элементы строения его частей. Лист снаружи покрыт живой тканью кожицей, в которой находятся отверстия - устьица. Они необходимы для защиты и газообмена. Внутреннее содержимое листа представлено основной запасающей и хлорофиллоносной тканью, которая отвечает за автотрофное питание всего растения. Элементы проводящей и образуя жилки, являются основой для транспортировки всей гаммы необходимых питательных веществ.

Являются корень (его подземная часть) и побег. Строение генеративных частей не позволяет им выполнять подобные функции. Они осуществляют процесс полового размножения и распространения растений. Но цветок развивается именно на стебле, а для его развития необходимы органические вещества, образованные в листьях.
Можно сделать вывод, что растение является единым организмом, функционирование частей которого взаимосвязано.
Развертывание побега из почки и его развитие. В жизни побега различают 2 этапа. Период формирования побега как зачаточного образования называют внутрипочечным или эмбриональным . При развертывании почки эмбриональный период в жизни побега сменяются внепочечным или постэмбриональным периодом. С наступлением весны почки трогаются в рост и отрастают новые побеги (стебли с листьями и почками). Пронаблюдать процесс развертывания побега можно еще до наступления весны, поставив в воду ветки деревьев или кустарников (особенно во второй половине зимы). Развертывание побега начинается с набухания почки, почечные чешуи раздвигаются, зачатки зеленых листьев увеличиваются в размерах. Вскоре после прорастания почки почечные чешуи опадают, а оставшиеся рубцы от опавших чешуй образуют на побеге почечное кольцо. По длительно сохраняющимся почечным рубцам можно установить возраст ветви дерева или кустарника. Одновременно наблюдается удлинение междоузлий за счет активно делящихся клеток вставочной меристемы. В этот период происходит интенсивный рост листовых пластинок с морфологически верхней стороны, и лист отгибается от стебля. Благодаря вставочному росту между основанием листа и пластинкой формируется черешок. Закладка боковых побегов происходит как внутри материнской почки, так и в период внепочечного роста побега.
Понятие о годичном и элементарном побегах. Годичный побег – побег, рост и формирование которого во внепочечный период жизни заканчивается в течение одного года. В условиях сезонного климата это происходит за один вегетационный период. Интенсивность роста и развития отдельных метамеров различна. Часто у основания побега междоузлия короткие и узлы сближены, выше по побегу, они становятся более длинными, а на верхушке снова наблюдается уменьшение длины междоузлий (максимальные размеры междоузлий, листьев, почек соответствуют срединным метамерам). Развитие элементарного побега происходит по одновершинной кривой за один ростовой толчок, или один период видимого роста. Нередко у дуба при исследовании годичного побега можно отметить, что он сформировался в результате двух периодов роста. Таким образом, следует говорить, что годичный побег состоит из двух элементарных побегов. Между двумя элементарными побегами отсутствуют чешуевидные листья, то есть почечное кольцо не формируется. У растений бессезонного климата годичные побеги состоят, как правило, из нескольких элементарных побегов.
Морфологические типы побегов. Побеги различаются:
1. По длине междоузлий. Удлиненный – побег, у которого четко выражены междоузлия и узлы находятся далеко друг от друга. Укороченные – побеги, у которых узлы сближены, и междоузлия практически не выражены или отсутствуют (подорожник). У одного и того же растения наряду с удлиненными побегами могут развиваться и укороченные (яблоня, береза, осока волосистая). Обычно укороченные побеги характеризуются небольшим годичным приростом.
У некоторых растений (сосна, плаун) годичные побеги обычно более 10 см длины, но имеют сближенные междоузлия. Такие побеги лучше называть длинными (рис. 6). Укороченные побеги травянистых растений называют розеточными (примула, одуванчик, подорожник). Полурозеточные побеги (живучка ползучая, нивяник, василек луговой, колокольчик персиколистный) характеризуются сближенными узлами в базальной части побега и удлиненными – в средней его части. В области соцветия узлы могут быть как удлиненными (колокольчик раскидистый), так и сближенными (колокольчик скученный). У пастушьей сумки обыкновенной, редьки дикой и других по мере раскрывания цветков происходит удлинение узлов в соцветии.
2. По функциям. У многих растений наблюдается специализация побегов. У древесных растений удлиненные побеги часто вегетативные (выполняют ростовую и трофическую функцию), а укороченные – генеративные . У вяза, бобовника, волчьего лыка полностью отсутствуют зеленые листья на генеративных побегах. У травянистых растений часто наблюдается обратная корреляция. Укороченные побеги – вегетативные, а удлиненные – генеративные (ландыш, подорожник).
3. Положение побегов в пространстве. Побеги могут быть прямостоячими (или ортотропными), горизонтальными (или плагиотропными), приподнимающимися (или анизотропными), наклонными, обвивающимися вокруг опоры, цепляющимися за опору (рис. 4). Разнообразие положения побегов разных растений в пространстве позволяет произрастать большему числу видов на той или иной территории.
4. По времени формирования побегов из почек. Мы уже рассматривали (см. Разнообразие почек) особенности формирования побегов обогащения (силлептические), побегов возобновления и водяных побегов.
Образование системы побегов. Формирование системы побегов происходит благодаря их ветвления и нарастания. Ветвление побега – процесс, приводящий к формированию побега на материнском побеге, то есть на побеге одного порядка формируются побеги следующего порядка.
Существует два типа ветвления побега: 1) верхушечное, 2) боковое. В процессе исторического развития растений характер ветвления изменялся. Верхушечное, или дихотомическое ветвление характерно для плаунов, некоторых папоротников и отдельных семенных растений (некоторых пальм). Выделяют равнодихотомическое (равновильчатое) ветвление – формирующиеся побеги одинаковые и неравнодихотомичское (неравновильчатое) – один побег оказывается более мощным и является как бы продолжением материнского побега.
Рис. 4. Типы побегов по расположению в пространстве: 1) – ортотропный (гвоздика Фишера); 2) – плагиотропный (вербейник монетчатый); 3) – анизотропный (плаун); 4) – вьющийся (вьюнок полевой); 5) – цепляющийся (горошек мышиный); 6) – наклонный (береза повислая)
Для подавляющего большинства семенных растений характерно боковое ветвление. Начало новому побегу дают боковые почки. Образование боковых побегов увеличивает их общее число. Возрастает общая поверхность органов воздушного питания, что чрезвычайно важно для растений, ведущих «прикрепленный» образ жизни.
За счет верхушечной почки происходит нарастание побега в длину. В течение ряда лет в результате деятельности одной меристемы формируются многолетние оси одного порядка. Такой тип нарастания называется моноподиальным (рис. 5). Так нарастает клен, ель и другие. Однако у ряда растений верхушечная меристема на определенном этапе формирует соцветие и дальнейшее моноподиальное нарастание становится невозможным.
У некоторых видов (береза, ива, липа) происходит отмирание верхушечной почки или даже части побега. Нарастание подобных растений происходит из боковых почек. Осенью после отмирания верхушечной почки и части побега одна из боковых почек становится по положению верхушечной, но наличие рубца от отмершей верхушки побега (веточного рубца) свидетельствует, что данная почка являются боковой. Такой тип нарастания называется симподиальным .
Таким образом, для семенных растений характерно два способа формирования многолетних осей: 1) моноподиальное нарастании и боковое ветвление (клен, дуб, ясень), 2) симподиальное нарастание и боковое ветвление (береза, ива, липа).
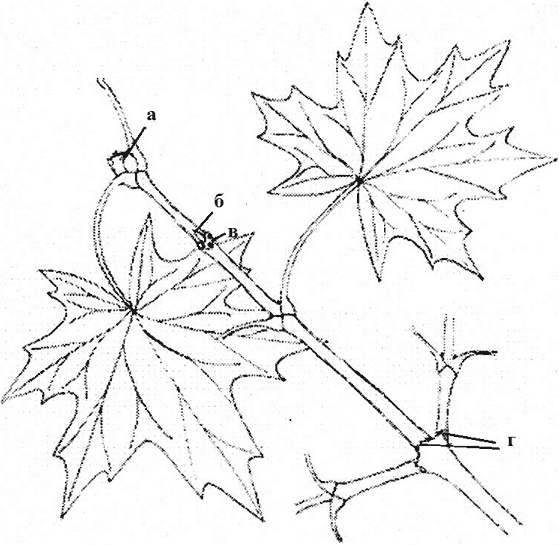
Рис. 5. Моноподиальное нарастание многолетних осей при боковом ветвлении. Ветка клена американского (2 года): а – верхушечная почка; б – пазушная почка; в – листовой рубец; г – почечное кольцо
Главный и боковые побеги. Главный побег формируется при прорастании семян из зародышевой почки. Из него в тот же год (или в последующие) начинает формироваться система побегов. Из верхушечной почки происходит нарастание главного побега, побега первого порядка. Из боковых почек в результате бокового ветвления формируются боковые побеги, побеги второго порядка. Боковые побеги второго также нарастают и ветвятся, образуя побеги третьего порядка, и так далее.
Акротония, мезотония, базитония. Эти три варианта ветвления побега различают в зависимости от расположения наиболее сильно развитых боковых побегов на материнском. При акротонном (греч. акрос – верхушка, тонос – сила, мощь) ветвлении наиболее мощные боковые побеги образуются на верхушке материнского побега, при мезотонном (греч. мезос – середина) – в середине, а при базитонном (греч. базис – основание) – в его основании. Частным случаем бокового ветвления является кущение побега. При этом боковые побеги формируются из почек, расположенных на укороченной части в основании побега.
Формирование ствола и кроны у деревьев. Деревья характеризуются образованием единственного ствола, обычно в его верхней части наблюдается интенсивное ветвление (акротонное), что приводит к формированию кроны. Нарастание ствола может быть как моноподиальным, так и симподиальным. В последнем случае ствол формируется в результате деятельности боковых почек по происхождению. Верхушечные почки, а чаще и небольшая верхняя часть побега развиты слабо и быстро отмирают. Формирование кроны происходит за счет пазушных почек и связано с разной интенсивностью ветвления. Угол наклона боковых ветвей относительно ствола также существенно влияет на своеобразие формы кроны. Обычно первые боковые ветви более слабые и быстро отмирают. Так у ели формирование полноценных ветвей кроны начинается только с 6–8 года, а иногда и позднее. Часто форма кроны напрямую зависит от условий произрастания растения. Одиноко стоящие деревья имеют значительно менее развитый ствол и более мощную крону. В густом лесу деревья образуют высокий ствол и небольшую крону на самой верхушке.
Формирование кустарников. Кустарники формируют несколько стволиков, которые сменяют друг друга по мере старения. Образование новых стволиков происходит за счет спящих почек, расположенных у основания материнского стволика. Они могут располагаться и приземно и подземно. Нарастание ствола происходит в течение нескольких лет. Ветвление происходит за счет пазушных почек. Степень ветвления различна у разных видов и часто зависит от фитоценоза. Если общая длительность жизни кустарника может достигать несколько сотен лет, то стволики живут около 20–40 лет. Однако эта величина колеблется в широких пределах: от 2 – у малины до 60 – у караганы.
Образование системы побегов у трав. Травянистые растения характеризуются большим разнообразием побеговых систем, которые образуются в результате бокового ветвления и моноподиального или симподиального нарастания. Обычно большая часть годичного прироста трав отмирает в год формирования. Многолетние побеговые системы обычно расположены в почве или плотно к ней прижаты. Наибольшее значение при характеристике побеговых систем травянистых растений имеет тип нарастания и длина годичного прироста. На основании этих признаков выделены модели побегообразования многолетних трав (длиннопобегая симподиальная – вероника длиннолистная, купена лекарственная; полурозеточная симподиальная – вереника колосистая, колокольчик персиколистный; розеточная моноподиальная – подорожник большой, одуванчик лекарственный; длиннопобеговая моноподиальная – вербейник монетчатый, вероника лекарственная).
Понятие о монокарпическом побеге. Монокарпический (моно – один, карпос – плод) побег цветет и плодоносит один раз. Понятие монокарпический побег, обычно используют для характеристики травянистых растений. Судьба монокарпического побега у разных растений может складываться по-разному:
1. Побег переходящий к цветению в первый год своего развития – моноциклический (купена многоцветковая, копытень европейский).
2. Побег, переходящий к цветению только на второй год жизни – дициклический побег (медуница неясная, земляника лесная, лютик кашубский).
3. Если побег переходит к цветению только на третий или в последующие годы – полициклический побег (грушанка круглолистная, овсяница овечья).
Кроме вышеназванных, существуют побеги, которые никогда не переходят к цветению. Они получили название побегов с неполным циклом развития. Причины этого могут быть разными: 1) неблагоприятные условия; 2) возрастное состояние; 3) специализация побегов у одного растения. К последней группе растений можно отнести моноподиально нарастающие побеги у подорожника большого, одуванчика лекарственного.
Листорасположение – это порядок расположения листьев на стебле (рис. 6). У одних растений от узла отходит лишь один лист, как, например, у березы, дуба, липы, лютика. Такое листорасположение называют очередным. Если на узле более одного листа – мутовчатое, его частным случаем является супротивное, при котором в пределах узла находятся два листа, обычно расположенных друг против друга (супротивно), как у клена, бузины, калины, вероники. У ряда видов (вороний глаз, ветреница, элодея, можжевельник) от узла отходят три или большее число листьев. Во всех случаях листья, отходящие от двух соседних узлов, никогда не располагаются друг над другом, а только под углом друг к другу. При таком листорасположении достигается минимальное затенение одного листа другим. Нередко у растений наблюдается неодинаковое разрастание черешков и пластинок и размещение листьев в одной плоскости, при этом образуется как бы сплошной зеленый экран, воспринимающий падающие лучи Солнца. Такое расположение листьев по отношению к источнику света (часто в условиях затенения) называют листовой мозаикой.
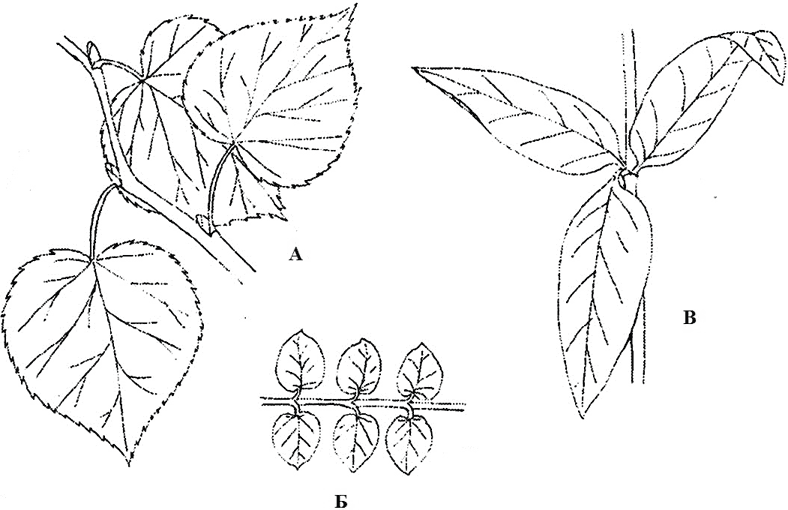
Рис. 6. Типы листорасположения: А – очередное (липа); Б – супротивное (вербейник монетчатый); В – мутовчатое (вербейник обыкновенный)
Стебель. Центральной, осьевой частью побега является стебель. Стебель выполняет опорную, транспортную и запасающую функции. Зеленые стебли также участвуют в воздушном питании растений. Стебель – опора для листьев, цветков, плодов, почек и развивающихся из них боковых побегов. По проводящим тканям стебля снизу вверх и сверху вниз осуществляется передача воды и растворенных в ней питательных веществ. В тканях стебля откладываются запасные вещества. Молодые зеленые стебли наряду с листьями участвуют в синтезе органических веществ из неорганических. У некоторых растений зеленые листья отсутствуют (саксаул, кактус, спаржа, иглица и другие), и стебель является основным органом воздушного питания.
На стебле выделяются узлы и междоузлия. Форму стебля обычно определяют по его поперечному срезу, сделанному на уровне междоузлия. У разных растений она не одинаковая, но постоянна для вида или даже рода, семейства. Это нередко имеет таксономическое значение. Чаще стебель округлый с ровным или ребристым краем. Он может быть четырехгранным (крапива, шалфей), трехгранным (осока), крылатым (чина лесная) и т. д. Стебель бывает гладким или опушенным, что определяется наличием различных волосков на эпидермисе.
Лист представляет собой боковой орган побега, расположенный на стебле. Функции листа: 1) фотосинтез, 2) транспирация, 3) газообмен. Более подробно с этими понятиями Вы познакомитесь в курсе физиологии растений.
Основными частями листа являются пластинка, черешок, прилистники и основание (рис. 7). Их строение соответствует тем функциям, которые лист выполняет, тем не менее, у разных растений они неодинаковые по форме и размерам (рис. 8). Пластинка – это расширенная, пластинчатая часть листа. Именно эта часть листа выполняет перечисленные выше функции. При пластинчатой форме органа достигается максимальная его поверхность, и как следствие высокая фотосинтетическая активность. В основании пластинка переходит в стеблевидный черешок. Его основная функция – размещение листовой пластинки в наиболее выгодном для растения положении в пространстве, а так же обеспечении пружинистости листа, то есть предотвращении повреждений листа при различных ударах. Он в свою очередь в нижней части переходит в основание листа, которое непосредственно связано со стеблем. Основание – это обязательная часть листа. У некоторых растений (морковь, пшеница) она разрастается и охватывает стебель над узлом. Такое основание называют влагалищем .
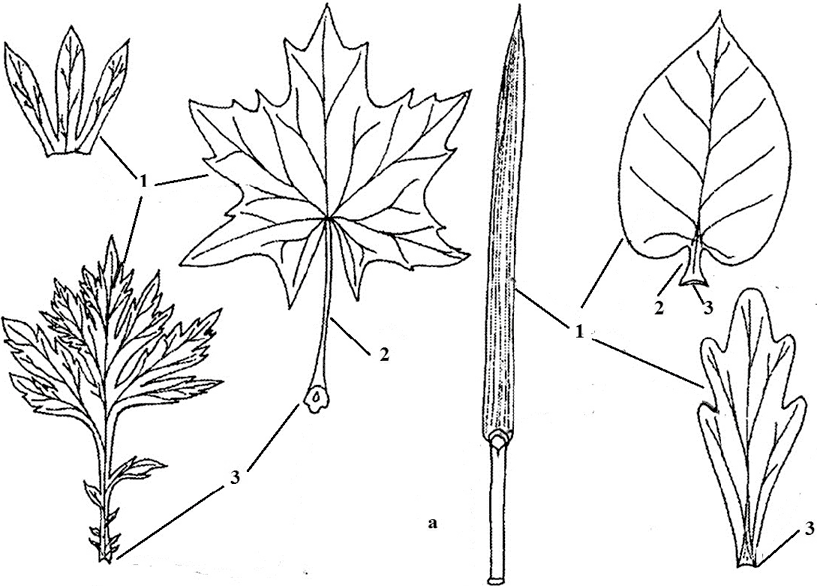
Рис. 7. Простые листья: 1 – листовая пластинка; 2 – черешок; 3 – основание; 4 – прилистники; а – влагалище; б – язычок; в – раструб
Прилистники – это выросты на основании листа. Их функция, в основном, связана с защитой листовой пластинки в период внутрипочечного развития. Однако у некоторых растений прилистники способны выполнять самостоятельные функции и во взрослом состоянии. Они могут значительно разрастаться как у гороха и напоминать пластинку, выполняя при этом фотосинтезурующую функцию. У желтой акации, крыжовника прилистники превращаются в колючки и служат как защитные образования. Лист называют полным, если у него есть пластинка, черешок, основание, прилистники. Полный лист у рябины, розы, дуба, черемухи. У первых двух из указанных растений все части листа сохраняются в течение всей жизни. У дуба взрослые листья не имеют прилистников, так как они рано отмирают, выполнив функции по защите пластинки зачаточного листа почки. При развертывании почки и формировании побега прилистники у дуба, березы, липы и ряда других растений опадают.

Рис. 8. Особенности морфологии листа: 1 – части листа: а – полный лист; б – сложный лист с прилистниками; в – прилистники срослись с черешком; г – раструб; д – ложномутовчатое листорасположение (например, у подмаренников), е – вздутое влагалище, ж – трубчатое влагалище (например, у злаков); 2 – положение листа на стебле: а – длинночерешковый лист, б – короткочерешковый, в – сидячий, г – низбегающий, д – стеблеобъемлющий, е – пронзенный, ж – сросшиеся листья; 3 – форма основания листовой пластинки: а – клиновидное, б – округлое, в – сердцевидное, г – срезанное, д – стреловидное, е – копьевидное, ж – неравнобокое, з – суженное; 4 – форма верхушки листа: а – тупая, б – усеченная, в – острая, г – заостренная, д – остроконечная, е – выемчатая; 5 – форма края листа: а – цельнокрайний, б – зубчатый, в – двоякозубчатый, г – пальчатый, д – двоякопильчатый, е – неравнопильчатый, ж – городчатый, з – выемчатый, и – волнистый, к – реснитчатый
Лист называют неполным, если у него отсутствует, хотя бы одна из его частей: черешок (лист сидячий), прилистники или пластинка. Сидячий лист у алоэ, колокольчика персиколистного, гвоздики Фишера. У этих растений так же отсутствуют и прилистники. Последних нет и у сирени, капусты, картофеля. Редко, но может отсутствовать пластинка. Тогда ее функции выполняют другие части: прилистники (чина безлисточковая), уплощенный черешок (у некоторых акаций).
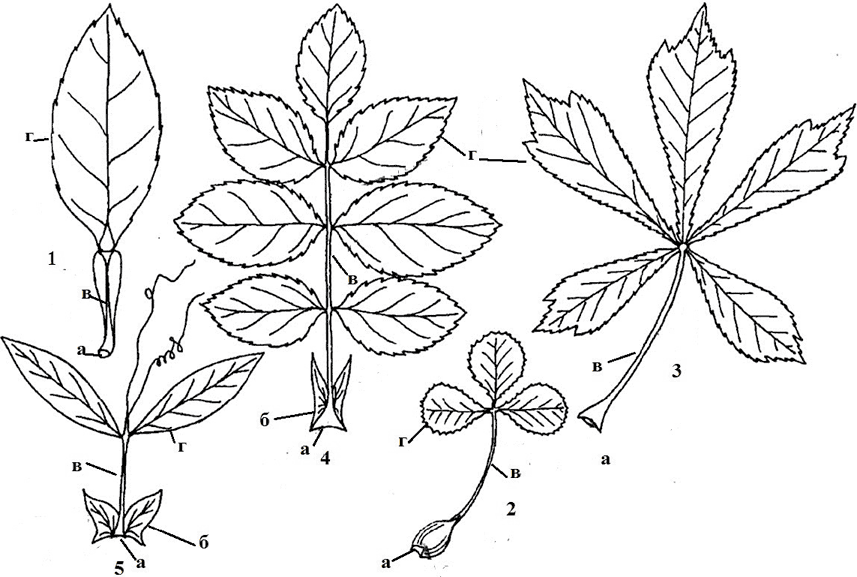
Рис. 9. Сложные листья: 1 – однолисточковый (лимон); 2 – тройчатосложный (название вида); 3 – пальчатый (каштан конский); 4 – парноперестый (чина луговая); 5 – непарноперестый (малина лесная); а – основание; б – прилистники; в – рахис; г – листочек; д – черешочек; е – прилистничек
Простые и сложные листья. Лист с одной пластинкой, не имеющий сочленения с черешком или основанием называют простым . Лист называют сложным (рис. 9), если у него одна или несколько пластинок, каждая из них имеет собственное сочленена с общим черешком – рахисом. Каждую листовую пластинку сложного листа называют листочком, или пластиночкой.
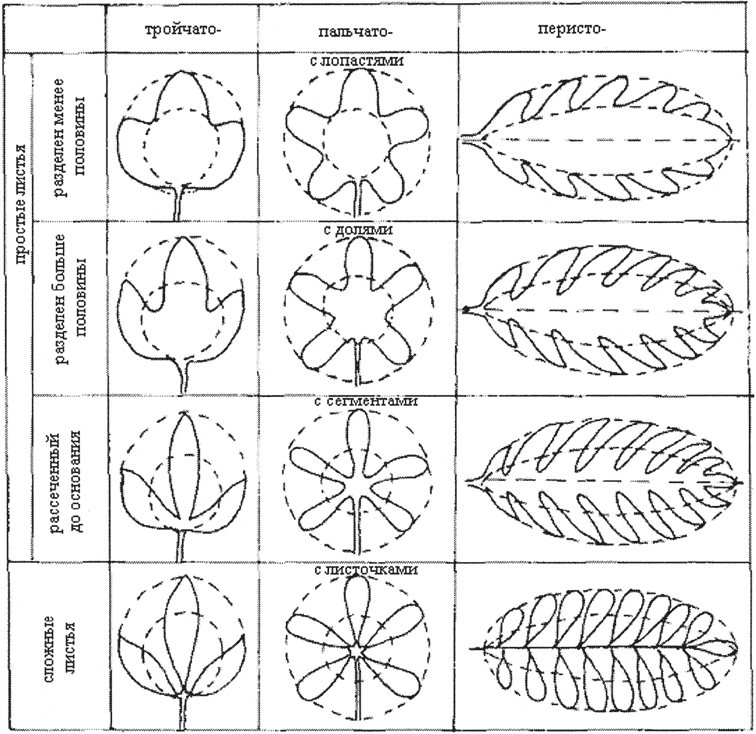
Рис. 10. Типы расчленения листа
Однолисточковый сложный лист – у лимона, мандарина, трехлисточковый сложный лист – у земляники, клевера, пальчатый лист – у люпина, каштана конского, непарноперистый лист – у рябины, ясеня (верхний листочек один, и только боковые листочки располагаются парами на общем черешке) и парноперистый – у сочевичника, гороха (все листочки занимают боковое положение на общем черешке и располагаются парами).
Сложные листья часто путают с простыми (рис. 10, 11), имеющими глубоко рассеченную пластинку: тройчаторассеченную – у ветреницы, пальчоторассеченную – у лапчатки прямостоячей, непарноперисторассеченную – у лапчатки гусиной, лировидный лист – у картофеля (лист непарноперисторассеченный с наиболее крупным верхним сегментом). Каждую отдельную часть пластинки называют сегментом. Сегмент не имеет сочленения с черешком. Форма и размеры листьев важный таксономический признак.
Разнообразие листовых пластинок. Пластинки простых листьев и листочки сложных листьев очень разнообразны по общему очертанию (округлые, овальные, яйцевидные, линейные и другие), по форме края пластинки (край может быть цельным, зубчатым, пильчатым, волнистым), характеру жилкования (рис. 11).
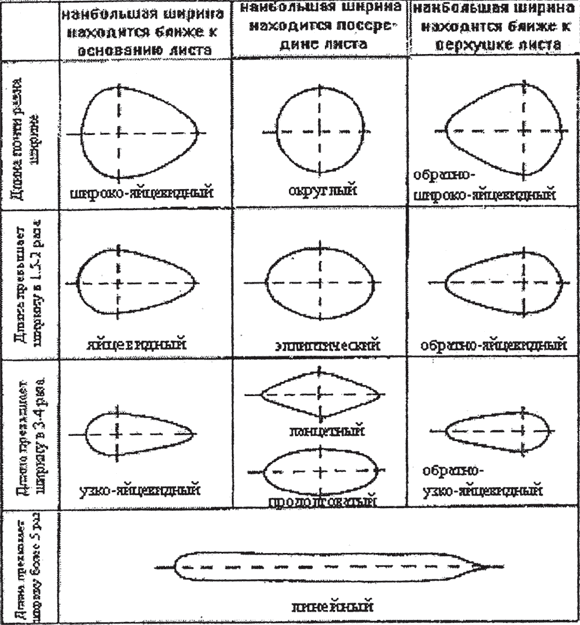
Рис. 11. Формы листовой пластинки
Многочисленные жилки пересекают пластинку в разных направлениях. На пластинке может быть одна мощная жилка, идущая по ее середине. Это главная жилка . От нее в стороны отходят более тонкие боковые, которые в свою очередь неоднократно ветвятся (береза, дуб). Такое жилкование пластинки называют перистым (или перисто-сетчатым). При наличии нескольких крупных, более или менее одинаковых жилок, сближенных в основании пластинки и расходящиеся веером (герань, лютик), жилкование называют пальчатым (или пальчато-сетчатым). Если крупные жилки проходят вдоль пластинки параллельно друг другу, то жилкование называют параллельным (пшеница, овсяница). Дуговидное жилкование наблюдается у листьев (ландыш, подорожник), крупные жилки, помимо центральной, изогнуты подобно дуге (рис. 12).
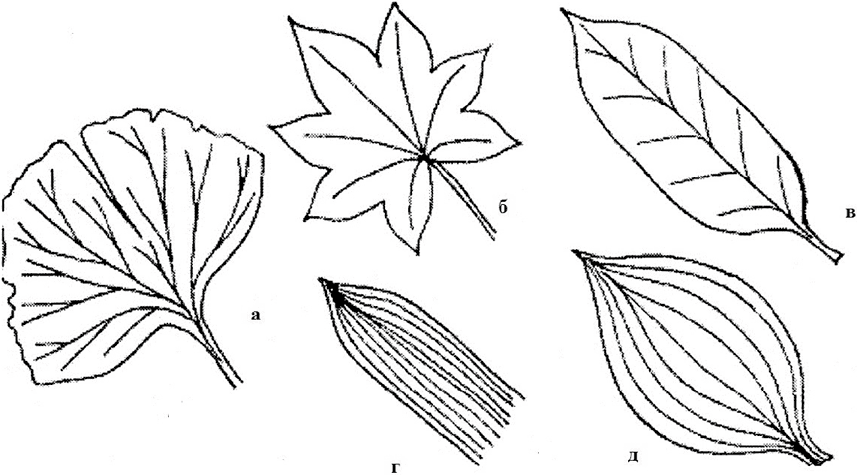
Рис. 12. Формы жилкования листа: а – дихотомическое; б – пальчатое; в – перистое; г – параллельное; д – дуговое
Три формации листьев. В основании годичного побега расположены листья низовой формации (почечные чешуи, чешуи луковиц), выполняющие защитную функцию. Обычно они чешуевидные или пленчатые, бурые, бледно-зеленые. Обычные зеленые листья образуют срединную формацию. Листья верховой формации расположены в области соцветия, являются кроющими листьями цветков и выполняют защитную функцию (для бутонов). У некоторых растений (марьянник дубравный) они окрашены в яркий цвет и служат для привлечения насекомых.
Побег , как и корень, - основной орган растения. Вегетативные побеги в типичном случае выполняют функцию воздушного питания, но имеют ряд других функций и способны к разнообразным метаморфозам. Спороносные побеги (в том числе и цветок) специализированы как органы репродуктивные , обеспечивающие размножение.
Побег образуется верхушечной меристемой как единое целое и, поэтому, представляет собой единый орган того же ранга, что и корень. Однако по сравнению с корнем побег имеет более сложное строение. Вегетативный побег состоит из осевой части – стебля , имеющего цилиндрическую форму, и листьев – плоских боковых органов, сидящих на стебле. Кроме того, обязательной частью побега являются почки – зачатки новых побегов, обеспечивающие нарастание побега и его ветвление, т.е. образование системы побегов. Главную функцию побега – фотосинтез – осуществляют листья; стебли – преимущественно несущие органы, выполняющие механическую и проводящую функции.
Главная черта, отличающая побег от корня, - его облиственность. Участок стебля, от которого отходит лист (листья) называется узел . Участки стебля между соседними узлами – междоузлия . Узлы и междоузлия повторяются вдоль оси побега. Таким образом, побег имеет метамерное строение, метамером (повторяющимся элементом) побега являются узел с листом и пазушной почкой и нижележащее междоузлие ( рис. 4.16).
Рис. 4.16. Строение побега.
Первый побег растения – его главный побег, или побег первого порядка. Он образуется из зародышевого побега, заканчивающегося почечкой , которая формирует все последующие метамеры главного побега. По положению эта почка – верхушечная ; пока она сохраняется, данный побег способен к дальнейшему росту в длину с образованием новых метамеров. Кроме верхушечной, на побеге образуются боковые почки. У семенных растений они находятся в пазухах листьев и называются пазушными . Из боковых пазушных почек развиваются боковые побеги, и происходит ветвление, за счет которого увеличивается общая фотосинтезирующая поверхность растения. Формируется система побегов , представленная главным побегом (побегом первого порядка) и боковыми (побегами второго порядка), а при повторении ветвления – боковыми побегами третьего, четвертого и последующих порядков. Побег любого порядка имеет свою верхушечную почку и способен к нарастанию в длину.
Почка – это зачаточный, еще не развернувшийся побег. Внутри почки заключена меристематическая верхушка побега – его апекс (рис. 4.17). Апекс представляет собой активно работающий ростовой центр, который обеспечивает формирование всех органов и первичных тканей побега. Источником постоянного самообновления апекса являются инициальные клетки апикальной меристемы, сосредоточенные на кончике апекса. Вегетативный апекс побега, в отличие от всегда гладкого апекса корня, регулярно формирует на поверхности выступы, представляющие собой зачатки листьев. Гладким остается лишь самый кончик апекса, который называют конусом нарастания побега. Его форма сильно варьирует у разных растений и не всегда имеет вид конуса, вершинная часть апекса может быть низкой, полушаровидной, плоской или даже вогнутой.
Из вегетативных почек развиваются вегетативные побеги, состоящие из стебля, листьев и почек. Такая почка состоит из меристематической зачаточной оси, оканчивающейся конусом нарастания , и зачаточных листьев разного возраста. Нижние листовые зачатки вследствие неравномерного роста загибаются внутрь и прикрывают верхние, более молодые, зачатки листьев и конус нарастания. Узлы в почке сближены, так как междоузлия еще не успели вытянуться. В пазухах листовых зачатков в почке уже могут быть заложены зачатки пазушных почек следующего порядка (рис. 4.17 ). В вегетативно-генеративных почках заложен ряд вегетативных метамеров, а конус нарастания превращен в зачаточный цветок или соцветие. Генеративные , или цветочные почки заключают в себе только зачаток соцветия или одиночного цветка, в последнем случае почку называют бутоном .
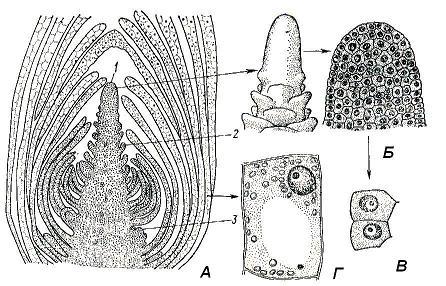
Рис. 4.17. Верхушечная почка побега элодеи: А – продольный срез; Б – конус нарастания (внешний вид и продольный срез); В – клетки апикальной меристемы; Г – паренхимная клетка сформировавшегося листа; 1 – конус нарастания; 2 – зачаток листа; 3 – зачаток пазушной почки.
Наружные листья почки часто видоизменяются в почечные чешуи , выполняющие защитную функцию и предохраняющие меристематические части почки от высыхания и резких перепадов температур. Такие почки называются закрытыми (зимующие почки деревьев и кустарников и некоторых многолетних трав). Открытые почки не имеют почечных чешуй.
Кроме обычных, экзогенных по заложению, пазушных почек у растений часто образуются придаточные , или адвентивные почки. Они возникают не в меристематической верхушке побега, а на взрослой, уже дифференцированной части органа эндогенно, из внутренних тканей. Придаточные почки могут образоваться на стеблях (тогда они обычно расположены в междоузлиях), листьях и корнях. Придаточные почки имеют большое биологическое значение: они обеспечивают активное вегетативное возобновление и размножение тех многолетних растений, у которых они имеются. В частности, при помощи придаточных почек возобновляются и размножаются корнеотпрысковые растения (малина, осина, осот, одуванчик). Корневые отпрыски – это побеги, развившиеся из придаточных почек на корнях. Придаточные почки на листьях образуются относительно редко. Если такие почки сразу дают маленькие побеги с придаточными корешками, которые отпадают от материнского листа и вырастают в новые особи, их называют выводковыми (бриофиллум).
В сезонном климате умеренной зоны развертывание побегов из почек у большинства растений носит периодический характер. У деревьев и кустарников, а также у многих многолетних травянистых растений почки разворачиваются в побеги один раз в году – весной или в начале лета, после чего формируются новые зимующие почки с зачатками побегов будущего года. Побеги, вырастающие из почек за один вегетационный период, называют годичными побегами , или годичными приростами . У деревьев они хорошо различаются благодаря образованию почечных колец – рубцов, которые остаются на стебле после опадения почечных чешуй. Летом у наших листопадных деревьев покрыты листьями годичные побеги только текущего года; на годичных побегах прошлых лет листьев уже нет. У вечнозеленых деревьев листья могут сохраняться на соответствующих годичных приростах 3-5 прошлых лет. В бессезонном климате за один год могут образоваться несколько побегов, разделенных небольшими периодами покоя. Такие побеги, образующиеся за один цикл роста, называют элементарными побегами .
Почки, впадающие на некоторое время в состояние покоя, а затем дающие новые элементарные и годичные побеги, называют зимующими или покоящимися . По функции их можно назвать почками регулярного возобновления . Такие почки – обязательный признак любого многолетнего растения, древесного или травянистого, именно они обеспечивают многолетность существования особи. По происхождению почки возобновления могут быть и экзогенными (верхушечными или пазушными), и эндогенными (придаточными).
Если боковые почки не имеют периода ростового покоя и развертываются одновременно с ростом материнского побега, их называют почками обогащения . Развертывающиеся из них побеги обогащения сильно увеличивают (обогащают) общую фотосинтезирующую поверхность растения, а также общее число образуемых соцветий и, следовательно, семенную продуктивность. Побеги обогащения характерны для большинства однолетних трав и для ряда многолетних травянистых растений с удлиненными цветоносными побегами.
Особую категорию составляют спящие почки , очень характерные для лиственных деревьев, кустарников, кустарничков и ряда многолетних трав. По происхождению они, как и почки регулярного возобновления, могут быть пазушными и придаточными, но, в отличие от них, не разворачиваются в побеги в течение многих лет. Стимулом для пробуждения спящих почек служит обычно или повреждение основного ствола или ветви (пневая поросль после порубки у ряда деревьев), или естественное старение материнской системы побегов, связанное с затуханием жизнедеятельности нормальных почек возобновления (смена стволиков у кустарников). У некоторых растений из спящих почек на стволе образуются безлистные цветоносные побеги. Это явление носит название каулифлории и характерно для многих деревьев тропических лесов, например для шоколадного дерева. У гледичии из спящих почек на стволе вырастают пучки крупных разветвленных колючек – видоизмененных побегов ( рис. 4.18).
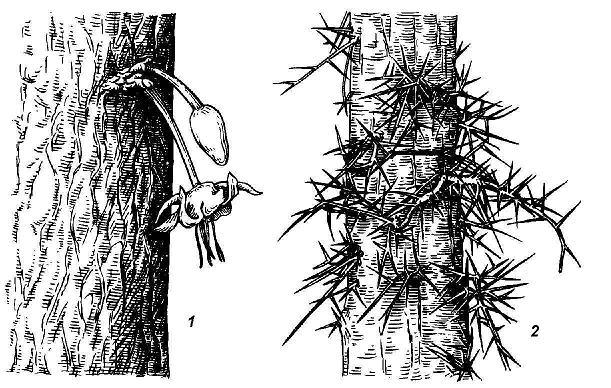
Рис. 4.18. Побеги из спящих почек: 1 – каулифлория у шоколадного дерева; 2 – колючки у гледичии из разветвившихся спящих почек.
Направление роста побегов. Побеги, растущие вертикально, перпендикулярно поверхности земли, носят название ортотропных . Горизонтально растущие побеги называются плагиотропными . Направление роста может меняться в процессе развития побега.
В зависимости от положения в пространстве различают морфологические типы побегов (рис. 4.19 ). Главный побег в большинстве случаев сохраняет ортотропный рост и остается прямостоячим . Боковые побеги могут расти в различном направлении, часто они образуют разной величины угол с материнским побегом. В процессе роста побег может изменить направление с плагиотропного на ортотропное, тогда он называется приподнимающимся , или восходящим . Побеги с сохраняющимся в течение жизни плагиотропным ростом называются стелющимися . Если они образуют в узлах придаточные корни, их называют ползучими .
Ортотропный рост связан определенным образом со степенью развития механических тканей. При отсутствии хорошо развитых механических тканей в удлиненных побегах ортотропный рост невозможен. Но нередко растения, не обладающие достаточно развитым внутренним скелетом, все же растут вверх. Это достигается различными путями. Слабые побеги таких растений - лиан закручиваются вокруг какой-либо твердой опоры (вьющиеся побеги), лазят с помощью различного рода шипиков, крючков, корней - прицепок (лазящие побеги), цепляются с помощью усиков различного происхождения (цепляющиеся побеги).
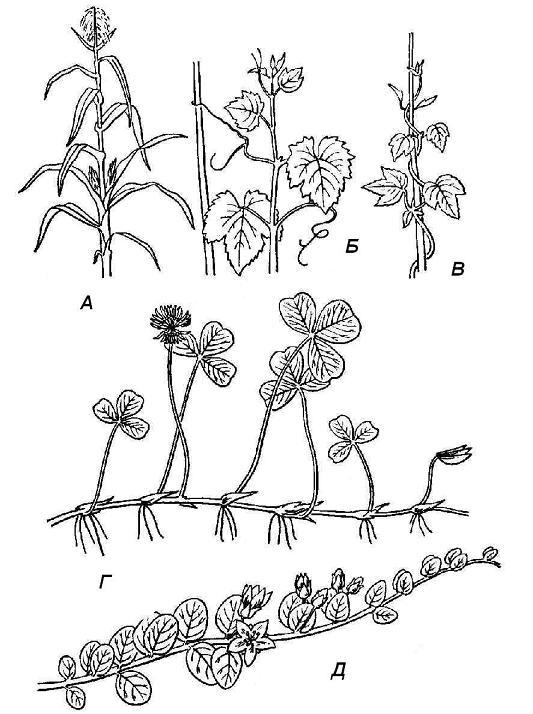
Рис. 4.19. Типы побегов по положению в пространстве : А – прямостоячий; Б – цепляющийся; В – вьющийся; Г – ползучий; Д – стелющийся.
Листорасположение. Листорасположение , или филлотаксис – порядок размещения листьев на оси побега. Различают несколько основных типов листорасположения (рис. 4.20 ).
Спиральное , или очередное листорасположение наблюдается, когда на каждом узле расположен один лист, и основания последовательных листьев можно соединить условной спиральной линией. Двурядное листорасположение можно рассматривать как частный случай спирального. При этом на каждом узле находится один лист, охватывающий широким основанием всю или почти всю окружность оси. Мутовчатое листорасположение возникает, если на одном узле закладывается несколько листьев. Супротивное листорасположение – частный случай мутовчатого, когда на одном узле образуются два листа, точно друг против друга; чаще всего такое листорасположение бывает накрест супротивным , т.е. соседние пары листьев находятся во взаимно перпендикулярных плоскостях ( рис. 4.20).
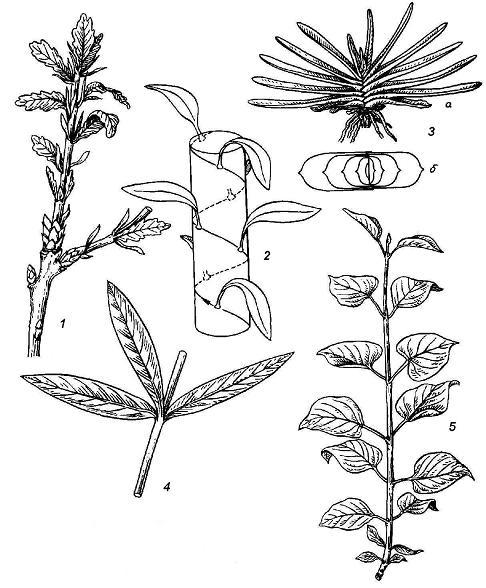
Рис. 4.20. Типы листорасположения : 1 – спиральное у дуба; 2 – схема спирального листорасположения; 3 – двурядное у гастерии (а – вид растения сбоку, б – вид сверху, схема); 4 – мутовчатое у олеандра; 5 – супротивное у сирени.
Порядок заложения листовых зачатков на апексе побега – наследственный признак каждого вида, иногда характерный для рода и даже целого семейства растений. Листорасположение взрослого побега определяется в первую очередь генетическими факторами. Однако в процессе развертывания побега из почки и его дальнейшего роста на расположение листьев могут оказывать влияние внешние факторы, главным образом, условия освещения и сила тяжести. Поэтому окончательная картина листорасположения может сильно отличаться от первоначальной и обычно приобретает ярко выраженный приспособительный характер. Листья располагаются так, что их пластинки оказываются в наиболее благоприятных в каждом конкретном случае условиях освещения. Наиболее ярко это проявляется в форме листовой мозаики , наблюдаемой на плагиотропных и розеточных побегах растений. При этом пластинки всех листьев располагаются горизонтально, листья не затеняют друг друга, а образуют единую плоскость, где нет просветов; более мелкие листья заполняют промежутки между крупными.
Типы ветвления побега. Ветвление – это образование системы осей. Оно обеспечивает увеличение общей площади соприкосновения тела растения с воздушной средой, водой или почвой. Ветвление возникло в процессе эволюции еще до появления органов. В простейшем случае верхушка главной оси вильчато ветвится и дает начало двум осям следующего порядка. Это верхушечное , или дихотомическое ветвление. Верхушечным ветвлением обладают многие многоклеточные водоросли, а также некоторые примитивные растения, например плауны (рис. 4.21 ).
Для остальных групп растений характерен более специализированный боковой тип ветвления. При этом боковые ветви закладываются ниже верхушки главной оси, не затрагивая ее способности к дальнейшему нарастанию. При таком способе потенциальные возможности ветвления и образования систем органов гораздо более обширны и биологически выгодны.
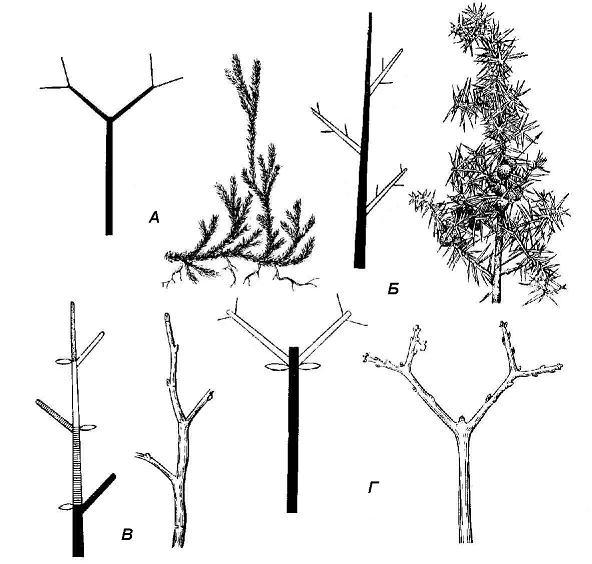
Рис. 4.21. Типы ветвления побега: А – дихотомическое (плаун); Б – моноподиальное (можжевельник); В – симподиальное по типу монохазия (черемуха); Г – симподиальное по типу дихазия (клен).
Различают два типа бокового ветвления: моноподиальное и симподиальное (рис. 4.21 ). При моноподиальной системе ветвления каждая ось представляет собой моноподий, т.е. результат работы одной верхушечной меристемы. Моноподиальное ветвление характерно для большинства голосеменных и многих травянистых покрытосеменных растений. Большинство покрытосеменных растений, однако, ветвится по симподиальному типу. При симподиальном ветвлении верхушечная почка побега на определенном этапе отмирает или прекращает активный рост, но начинается усиленное развитие одной или нескольких боковых почек. Из них образуются побеги, замещающие побег, прекративший рост. Образовавшаяся при этом ось представляет собой симподий – составную ось, состоящую из осей нескольких последовательных порядков. Способность растений к симподиальному ветвлению имеет большое биологическое значение. В случае повреждения верхушечной почки рост оси будет продолжен боковыми побегами.
В зависимости от числа замещающих осей различают симподиальное ветвление по типу монохазия , дихазия и плейохазия . Ветвление по типу дихазия, или ложнодихотомическое ветвление характерно для побегов с супротивным листорасположением (сирень, калина).
У некоторых групп растений нарастание основных скелетных осей идет за счет одной или немногих верхушечных почек, боковые скелетные ветви совсем не образуются или образуются в очень малом числе. Древовидные растения такого типа встречаются главным образом в тропических областях (пальмы, драцены, юкки, агавы, саговники). Крона этих растений образована не ветвями, а крупными листьями, сближенными в розетку на верхушке ствола. Способность к быстрому разрастанию и захвату пространства, а также к восстановлению после повреждений у таких растений нередко отсутствует или слабо выражена. Среди деревьев умеренного климата такие неветвящиеся формы практически не встречаются.
Другая крайность – растения, слишком обильно ветвящиеся. Они представлены жизненной формой растений-подушек (рис. 4.22 ). Нарастание в длину побегов у этих растений крайне ограничено, но зато ежегодно образуется множество боковых веточек, расходящихся во всех направлениях. Поверхность системы побегов растения выглядит как бы подстриженной; некоторые подушки настолько плотны, что похожи на камни.
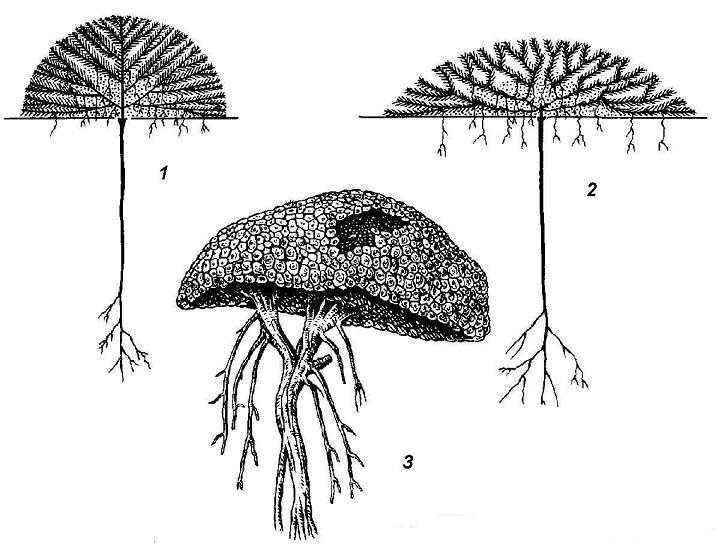
Рис. 4.22. Растения – подушки : 1, 2 – схемы строения растений-подушек; 3 – азорелла с острова Кергелен.
Очень сильно ветвятся представители жизненной формы перекати-поле , характерной для степных растений. Шаровидно разветвленная, очень рыхлая система побегов представляет собой огромное соцветие, которое после созревания плодов отламывается у основания стебля и перекатывается ветром по степи, рассеивая семена.
Специализация и метаморфозы побегов. У многих растений в пределах системы побегов наблюдается определенная специализация. Ортотропные и плагиотропные, удлиненные и укороченные побеги выполняют разные функции.
Удлиненными называют побеги с нормально развитыми междоузлиями. У древесных растений они называются ростовыми и располагаются по периферии кроны, определяя ее форму. Их главная функция – захват пространства, увеличение объема фотосинтезирующих органов. Укороченные побеги имеют сближенные узлы и очень короткие междоузлия (рис. 4.23 ). Они формируются внутри кроны и поглощают проникающий туда рассеянный свет. Часто укороченные побеги у деревьев являются цветоносными и выполняют функцию размножения.
Рис. 4.23. Укороченный (А) и удлиненный (Б) побеги платана: 1 – междоузлие; 2 – годичные приросты.
У травянистых растений обычно укороченные розеточные побеги выполняют функцию многолетних скелетных и фотосинтезирующих, а удлиненные формируются в пазухах розеточных листьев и являются цветоносными (подорожник, манжетка, фиалки). Если пазушные цветоносы безлистны, их называют стрелками . Тот факт, что у древесных растений цветоносные побеги укороченные, а у травянистых – удлиненные, биологически хорошо объясним. Для успешного опыления соцветия трав должны быть приподняты над травостоем, а у деревьев даже укороченные побеги в кроне находятся в благоприятных для опыления условиях.
Примером специализации побегов могут служить многолетние осевые органы древесных растений – стволы и сучья кроны. У листопадных деревьев годичные побеги теряют ассимиляционную функцию после первого вегетационного периода, у вечнозеленых – через несколько лет. Часть побегов после потери листьев отмирает целиком, но большинство остается в качестве скелетных осей, выполняющих опорную, проводящую и запасающую функции в течение десятков лет. Лишенные листьев скелетные оси известны под названием сучьев и стволов (у деревьев), стволиков (у кустарников).
В ходе приспособления к специфическим условиям среды обитания или в связи с резкой сменой функций побеги могут видоизменяться (метаморфизироваться). Особенно часто метаморфизируются побеги, развивающиеся под землей. Такие побеги утрачивают функцию фотосинтеза; они обычны у многолетних растений, где выступают в качестве органов переживания неблагоприятного периода года, запаса и возобновления.
Наиболее часто встречающимся подземным метаморфозом побега является корневище (рис. 4.24). Корневищем принято называть долговечный подземный побег, выполняющий функции отложения запасных питательных веществ, возобновления, а иногда и вегетативного размножения. Корневище формируется у многолетних растений, как правило, не имеющих во взрослом состоянии главного корня. По положению в пространстве оно может быть горизонтальным , косым или вертикальным . Корневище обычно не несет зеленых листьев, но, будучи побегом, сохраняет метамерную структуру. Узлы выделяются либо по листовым рубцам и остаткам сухих листьев, либо по живым чешуевидным листьям, также в узлах располагаются пазушные почки. По этим признакам корневище легко отличить от корня. Как правило, на корневище формируются придаточные корни; из почек вырастают боковые ответвления корневища и надземные побеги.
Формируется корневище или изначально как подземный орган (купена, вороний глаз, ландыш, черника), или сначала как надземный ассимилирующий побег, который затем погружается в почву с помощью втягивающих корней (земляника, медуница, манжетка). Корневища могут нарастать и ветвиться моноподиально (манжетка, вороний глаз) или симподиально (купена, медуница). В зависимости от длины междоузлий и интенсивности нарастания, различают длинные и короткие корневища и, соответственно, длиннокорневищные и короткокорневищные растения.
При ветвлении корневищ образуется куртина надземных побегов, связанных участками системы корневищ. Если связующие части разрушаются, побеги обособляются, и происходит вегетативное размножение. Совокупность новых особей, образовавшихся вегетативным путем, называют клоном . Корневища характерны преимущественно для травянистых многолетников, но встречаются и у кустарников (бересклет) и кустарничков (брусника, черника).
Близки к корневищам подземные столоны – недолговечные тонкие подземные побеги, несущие недоразвитые чешуевидные листья. Столоны служат для вегетативного размножения, расселения и захвата территории. Запасные питательные вещества в них не откладываются.
У некоторых растений (картофель, земляная груша) к концу лета из верхушечных почек столонов образуются клубни (рис. 4.24 ). Клубень имеет шаровидную или овальную форму, стебель сильно утолщен, в нем откладываются запасные питательные вещества, листья редуцируются, в их пазухах формируются почки. Столоны отмирают и разрушаются, клубни перезимовывают, и на следующий год дают начало новым надземным побегам.
Клубни не всегда развиваются на столонах. У некоторых многолетних растений клубневидно разрастается и утолщается основание главного побега (цикламен, капуста кольраби) (рис. 4.24 ). Функции клубня – запас питательных веществ, переживание неблагоприятного периода года, вегетативное возобновление и размножение.
У многолетних трав и полукустарничков с хорошо развитым стержневым корнем, сохраняющимся в течение всей жизни, образуется своеобразный орган побегового происхождения, называемый каудексом . Вместе с корнем он служит местом отложения запасных веществ и несет множество почек возобновления, часть из которых могут быть спящими. Каудекс обычно бывает подземным и образуется из коротких оснований побегов, погружающихся в почву. От коротких корневищ каудекс отличается способом отмирания. Корневища, нарастая верхушкой, постепенно отмирают и разрушаются на более старом конце; главный корень не сохраняется. Каудекс разрастается в ширину, с нижнего конца он постепенно переходит в долгоживущий утолщающийся корень. Отмирание и разрушение каудекса и корня идет от центра к периферии. В центре образуется полость, а затем он может разделиться продольно на отдельные участки – партикулы . Процесс разделения особи стержнекорневого растения с каудексом на части называют партикуляцией . Каудексовых растений много среди бобовых (люпин, люцерны), зонтичных (бедренец, ферула), сложноцветных (одуванчик, полыни).
Луковица – это, как правило, подземный побег с очень коротким уплощенным стеблем – донцем и чешуевидными мясистыми сочными листьями, запасающими воду и растворимые питательные вещества, главным образом сахара. Из верхушечной и пазушных почек луковиц вырастают надземные побеги, на донце образуются придаточные корни (рис. 4.24 ). Таким образом, луковица – типичный орган вегетативного возобновления и размножения. Луковицы наиболее характерны для растений из семейств лилейных (лилии, тюльпаны), луковых (луки) и амариллисовых (нарциссы, гиацинты).
По структуре луковицы очень разнообразны. В одних случаях запасающие чешуи луковицы представляют собой только видоизмененные листья, не имеющие зеленых пластинок (лилия саранка); в других – это подземные влагалища зеленых ассимилирующих листьев, которые утолщаются и остаются в составе луковицы после отмирания пластинок (лук репчатый). Нарастание оси луковицы может быть моноподиальным (подснежник) или симподиальным (гиацинт). Наружные чешуи луковицы расходуют запас питательных веществ, подсыхают и играют защитную роль. Число луковичных чешуй варьирует от одной (чеснок) до нескольких сотен (лилии).
В качестве органа возобновления и запаса луковица приспособлена главным образом к климатам средиземноморского типа – с достаточно мягкой влажной зимой и с очень жарким засушливым летом. Она служит не столько для благополучной перезимовки, сколько для переживания жесткой летней засухи. Запасание воды в тканях луковичных чешуй происходит за счет образования слизей, способных удерживать большое количество воды.
Клубнелуковица внешне напоминает луковицу, но ее чешуевидные листья не являются запасающими; они сухие и пленчатые, а запасные вещества откладываются в утолщенной стеблевой части (шафран, гладиолус).

Рис. 4.24. Подземные метаморфозы побега : 1, 2, 3, 4 – последовательность развития и строение клубня картофеля; 5 – клубень цикламена; 6 – клубень кольраби; 7 – луковички лилии тигровой; 8 – луковица лука репчатого; 9 – луковица лилии; 10 – участок длинного корневища пырея ползучего.
Видоизменяться могут не только подземные, но и надземные побеги растений (рис. 4.25 ). Довольно часто встречаются надземные столоны . Это плагиотропные недолговечные побеги, функция которых вегетативное размножение, расселение и захват территории. Если столоны несут зеленые листья и участвуют в процессе фотосинтеза, их называют плетями (костяника, живучка ползучая). У земляники столоны лишены развитых зеленых листьев, стебли у них тонкие и хрупкие, с очень длинными междоузлиями. Такие более высокоспециализированные для функции вегетативного размножения столоны называют усами .
Сочными, мясистыми, приспособленными для накопления воды могут быть не только луковицы, но и надземные побеги, обычно у растений, живущих в условиях недостатка влаги. Водозапасающими органами могут быть листья или стебли, иногда даже почки. Такие сочные растения носят название суккулентов . Листовые суккуленты запасают воду в тканях листьев (алоэ, агавы, толстянки, родиола, или золотой корень). Стеблевые суккуленты характерны для американского семейства кактусовых и африканских молочайных. Сочный стебель выполняет водозапасающую и ассимилирующую функцию; листья редуцированы или превращены в колючки (рис. 4.25, 1). У большинства кактусов стебли колонновидные или шаровидные, листьев на них совсем не образуется, но узлы хорошо заметны по расположению пазушных побегов – ареол , имеющих вид бородавок или удлиненных выростов с колючками или пучками волосков. Превращение листьев в колючки уменьшает испаряющую поверхность растения и защищает от поедания животными. Примером метаморфоза почки в суккулентный орган – кочан служит культурная капуста.
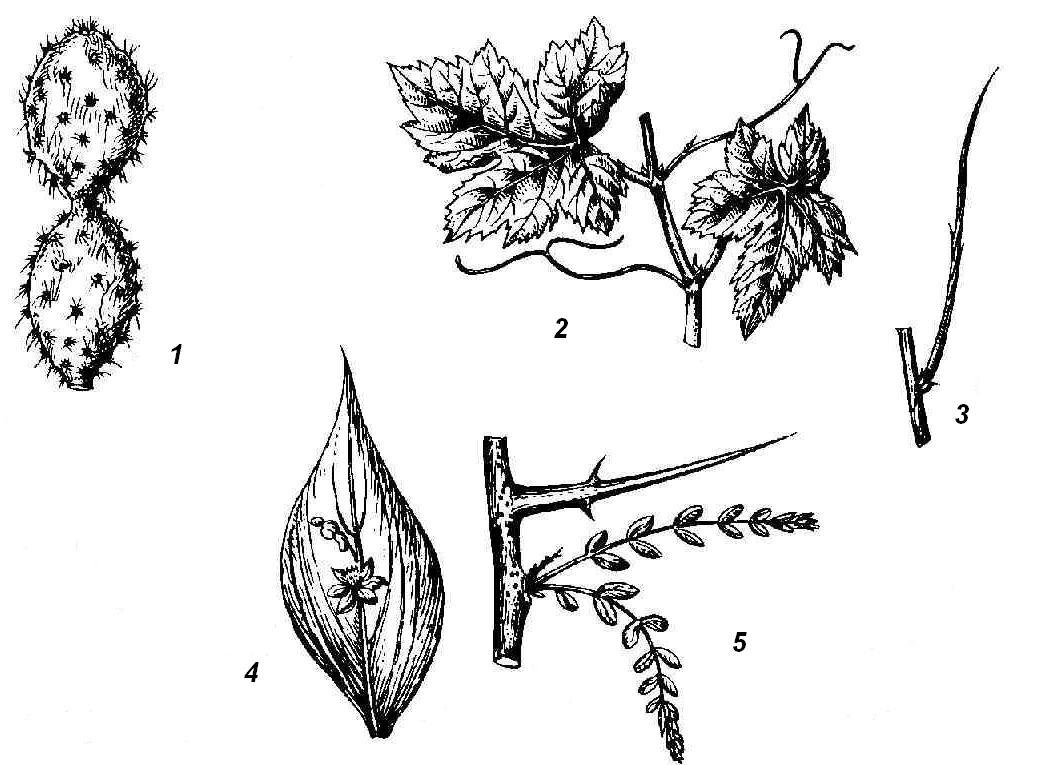
Рис. 4.25. Надземные метаморфозы побега: 1 – стеблевой суккулент (кактус); 2 – усики винограда; 3 – безлистный фотосинтезирующий побег дрока; 4 – филлокладий иглицы; 5 – колючка гледичии.
Колючки кактусов имеют листовое происхождение. Листовые колючки нередко встречаются и у несуккулентных растений (барбарис) (рис. 4.26, 1). У многих растений колючки имеют не листовое, а стеблевое происхождение. У дикой яблони, дикой груши, жостера слабительного в колючки метаморфизируются укороченные побеги, имеющие ограниченный рост и оканчивающиеся острием. Вид жесткой одревесневшей колючки они приобретают после опадения листьев. У боярышника (рис . 4.26, 3) колючки, образующиеся в пазухах листьев, совершенно безлистны с самого начала. У гледичии (рис. 4.25, 5 ) мощные разветвленные колючки образуются на стволах из спящих почек. Образование колючек любого происхождения, как правило, есть результат недостатка влаги. При выращивании многих колючих растений в искусственной влажной атмосфере они утрачивают колючки: вместо них вырастают нормальные листья (верблюжья колючка) или облиственные побеги (английский дрок).
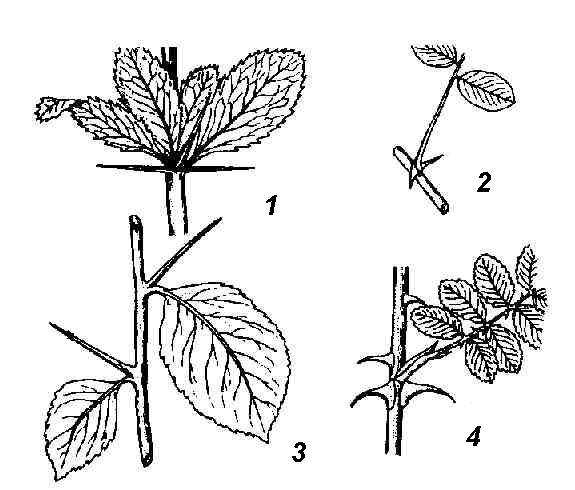
Рис. 4.26. Колючки различного происхождения : 1 – листовые колючки барбариса; 2 – колючки белой акации, видоизменение прилистников; 3 – колючки боярышника побегового происхождения; 4 – шипы – эмергенцы шиповника.
Побеги ряда растений несут шипы . Шипы отличаются от колючек меньшими размерами, это выросты – эмергенцы – покровной ткани и тканей коры стебля (шиповник, крыжовник) (рис. 4.26, 4 ).
Приспособление к недостатку влаги очень часто выражается в ранней утрате, метаморфозе или редукции листьев, теряющих основную функцию фотосинтеза. Это компенсируется тем, что роль ассимилирующего органа берет на себя стебель. Иногда такой ассимилирующий стебель безлистного побега остается внешне неизмененным (испанский дрок, верблюжья колючка) (рис. 4.25, 3). Дальнейшим шагом в указанной смене функций является формирование таких органов, как филлокладии и кладодии . Это уплощенные листоподобные стебли или целые побеги. На побегах иглицы (рис. 4.25, 4 ), в пазухах чешуевидных листьев, развиваются плоские листовидные филлокладии, имеющие, подобно листу, ограниченный рост. На филлокладиях образуются чешуевидные листья и соцветия, чего никогда не бывает на нормальных листьях, значит, филлокладий соответствует целому пазушному побегу. Мелкие, игольчатые филлокладии образуются у спаржи в пазухах чешуевидных листьев основного скелетного побега. Кладодии – это уплощенные стебли, сохраняющие, в отличие от филлокладиев, способность к длительному росту.
Для некоторых растений характерно видоизменение листьев или их частей, а иногда целых побегов в усики , которые закручиваются вокруг опоры, помогая тонкому и слабому стеблю сохранять вертикальное положение. У многих бобовых в усики превращается верхняя часть перистосложного листа (горох, горошки, чина). В других случаях в усики превращаются прилистники (сассапариль). Очень характерные усики листового происхождения формируются у тыквенных, причем можно видеть все переходы от нормальных листьев к полностью метаморфизированным. Усики побегового происхождения можно наблюдать у винограда (рис. 4.25, 2), пассифлоры и ряда других растений.