Паукообразные: строение, физиология и развитие
Класс паукообразных объединяет свыше 36 000 видов наземных хелицеровых, относящихся более чем к 10 отрядам.
Arachnida - высшие хелицеровые членистоногие с 6 парами головогрудных конечностей. Они дышат посредством легких или трахей и обладают помимо коксальных желез выделительным аппаратом в виде мальпигиевых сосудов, лежащих в брюшке.
Строение и физиология. Внешняя морфология. Тело паукообразных чаще всего состоит из головогруди и брюшка. В образовании головогруди участвуют акрон и 7 сегментов (7-й сегмент недоразвит). У сольпуг и некоторых других низших форм воедино спаяны лишь сегменты 4 передних пар конечностей, тогда как задние 2 сегмента головогруди свободны, а за ними следуют ясно разграниченные сегменты брюшка. Таким образом, у сольпуг имеются: передний отдел тела, по сегментарному составу отвечающий голове трилобитов (акрон + 4 сегмента), так называемый пропельтидий; два свободных грудных сегмента с ногами и сегментированное брюшко. Сольпуги, следовательно, принадлежат к паукообразным с наиболее богато расчлененным телом.
Следующий по степени расчлененности отряд - скорпионы , у которых головогрудь слитная, но за ней следует длинное 12-сегментное, как у Gigantostraca , брюшко, подразделяющееся на более широкое переднебрюшие (из 7 сегментов) и узкое заднебрюшие (из 5 сегментов). Тело заканчивается тельсоном, несущим искривленную ядовитую иглу. Таков же характер сегментации (только без подразделения брюшка на два участка) у представителей отрядов жгутоногих, лже-скорпионов, сенокосцев, у некоторых клещей и у примитивных членистобрюхих пауков.
Следующий этап слияния туловищных сегментов обнаруживают большинство пауков и некоторые клещи . У них не только головогрудь, но и брюшко представляют сплошные нерасчлененные отделы тела, однако у пауков между ними имеется короткий и узкий стебелек, образованный 7-м сегментом тела. Максимальная степень слияния сегментов тела наблюдается у ряда представителей отряда клещей, у которых все тело цельное, без границ между сегментами и без перетяжек.
Как уже говорилось, головогрудь несет 6 пар конечностей. Две передние пары участвуют в захвате и размельчении пищи - это хелицеры и педипальпы. Хелицеры располагаются впереди рта, чаще всего у паукообразных они в виде коротких клешней (сольпуги, скорпионы, лжескорпионы, сенокосцы, некоторые клещи и др.). Они обычно состоят из трех члеников, концевой членик играет роль подвижного пальца клешни. Реже хелицеры заканчиваются подвижным когтевидным члеником или имеют вид двучленистых придатков с заостренным и зазубренным краем, которым клещи прокалывают покровы животных.
Конечности второй пары, педипальпы, состоят из нескольких члеников. При помощи жевательного выроста на основном членике педипальп размельчается и разминается пища, тогда как прочие членики составляют род щупальца. У представителей некоторых отрядов (скорпионы, лжескорпионы) педипальпы превращены в мощные длинные клешни, у других - похожи на ходные ноги. Остальные 4 пары головогрудных конечностей состоят из 6-7 члеников и играют роль ходных ног. Они заканчиваются коготками.
У взрослых паукообразных брюшко лишено типичных конечностей, хотя они, несомненно, произошли от предков, обладавших хорошо развитыми ножками на передних брюшных сегментах. У эмбрионов многих паукообразных (скорпионов, пауков) на брюшке закладываются зачатки ножек, которые лишь впоследствии подвергаются регрессу. Впрочем, и во взрослом состоянии брюшные ножки иногда сохраняются, но в модифицированном виде. Так, у скорпионов на первом сегменте брюшка есть пара половых крышечек, под которыми открывается половое отверстие, на втором - пара гребенчатых органов, которые снабжены многочисленными нервными окончаниями и играют роль осязательных придатков. И те, и другие представляют видоизмененные конечности. Такова же природа и легочных мешков, расположенных на сегментах брюшка у скорпионов, некоторых пауков и лжескорпионов.
От конечностей берут свое начало и паутинные бородавки пауков. На нижней поверхности брюшка впереди порошицы у них находится 2-3 пары бугорков, усаженных волосками и несущих трубковидные протоки многочисленных паутинных желез. Гомология этих паутинных бородавок брюшным конечностям доказывается не только их эмбриональным развитием, но и строением их у некоторых тропических пауков, у которых бородавки особенно сильно развиты, состоят из нескольких члеников и даже по виду напоминают ножки.
Покровы хелицеровых состоят из кутикулы и подлежащих слоев: гиподермального эпителия (гиподермы) и базальной мембраны. Сама кутикула представляет сложное трехслойное образование. Снаружи располагается липопротеиновый слой, надежно предохраняющий организм от потери влаги при испарении. Это позволило хелицеровым стать настоящей сухопутной группой и заселить самые засушливые районы земного шара. Прочность кутикуле придают белки, задубленные фенолами и инкрустирующие хитин.
Производными кожного эпителия являются некоторые железистые образования, в том числе ядовитые и паутинные железы. Первые свойственны паукам, жгутоногим и скорпионам; вторые - паукам, лжескорпионам и некоторым клещам.
Пищеварительная система у представителей разных отрядов хелицеровых сильно варьирует. Передняя кишка обыкновенно образует расширение - снабженную сильными мышцами глотку, которая служит в качестве насоса, втягивающего полужидкую пищу, так как паукообразные не принимают твердую пищу кусками. В переднюю кишку открывается пара небольших "слюнных желез". У пауков секрет этих желез и печени способен энергично расщеплять белки. Он вводится в тело убитой добычи и приводит содержимое ее в состояние жидкой кашицы, которая затем всасывается пауком. Здесь имеет место так называемое внекишечное пищеварение.
У большинства паукообразных средняя кишка образует длинные боковые выпячивания, увеличивающие вместимость и всасывающую поверхность кишечника. Так, у пауков от головогрудной части средней кишки к основаниям конечностей идут 5 пар слепых железистых мешков; аналогичные выпячивания имеются у клещей, сенокосцев и других паукообразных. В брюшной отдел средней кишки открываются протоки парной пищеварительной железы - печени; она выделяет пищеварительные ферменты и служит для всасывания питательных веществ. В клетках печени происходит внутриклеточное пищеварение.
Выделительная система паукообразных по сравнению с мечехвостами имеет совершенно иной характер. На границе между средней и задней кишкой в пищеварительный канал открывается пара большей частью ветвящихся мальпигиевых сосудов. В отличие от Тгаcheata они энтодермального происхождения, т. е. образуются за счет средней кишки. Как в клетках, так и в просвете мальпигиевых сосудов есть многочисленные зерна гуанина - главного продукта выделения паукообразных. Гуанин, как и мочевая кислота, экскретируемая насекомыми, обладает малой растворимостью и удаляется из организма в виде кристаллов. Потери влаги при этом минимальны, что важно для животных, перешедших к жизни на суше.
Кроме мальпигиевых сосудов паукообразные обладают еще и типичными коксальными железами - парными мешковидными образованиями мезодермальной природы, лежащими в двух (реже в одном) сегментах головогруди. Они хорошо развиты у зародышей и в молодом возрасте, но у взрослых животных более или менее атрофируются. Вполне сформированные коксальные железы состоят из концевого эпителиального мешочка, петлевидно извитого канала и более прямого выводного протока с мочевым пузырьком и наружным отверстием. Концевой мешочек отвечает ресничной воронке целомодукта, отверстие которой замкнуто остатком целомического эпителия. Коксальные железы открываются у основания 3-й или 5-й пары конечностей.
Нервная система Arachnida разнообразна. Будучи связанной в происхождении с брюшной нервной цепочкой кольчатых червей, у паукообразных она обнаруживает четко выраженную тенденцию к концентрации.
Головной мозг имеет сложное строение. В его состав входят два отдела: передний, иннервирующий глаза, - протоцеребрум и задний - тритоцеребрум, посылающий нервы к первой паре конечностей - хелицерам. Характерный для других членистоногих (ракообразные, насекомые) промежуточный отдел мозга - дейтоцеребрум у паукообразных отсутствует. Связано это с исчезновением у них, как и у остальных хелицеровых, придатков акрона - антеннул, или усиков, которые иннервируются именно от дейтоцеребрума.
Метамерность брюшной нервной цепочки всего яснее сохраняется у скорпионов. У них имеется помимо головного мозга и окологлоточных коннективов большая ганглиозная масса в головогруди на брюшной стороне, дающая нервы к 2-6-й парам конечностей и 7 ганглиев, на протяжении брюшного отдела нервной цепочки. У сольпуг кроме сложного головогрудного ганглия на нервной цепочке сохраняется еще один узел, а у пауков уже вся цепочка слилась в головогрудный ганглий.
Наконец, у сенокосцев и клещей нет даже явственного разграничения между головным мозгом и головогрудным ганглием, так что нервная система образует вокруг пищевода сплошное ганглиозное кольцо.
Органы чувств Arachnida разнообразны. Очень важные для паукообразных механические, осязательные раздражения воспринимаются различно устроенными чувствительными волосками, которые особенно многочисленны на педипальпах. Специальные волоски - трихоботрии, расположенные на педипальпах, ногах и поверхности туловища, регистрируют колебания воздуха. Так называемые лировидные органы, представляющие собой небольшие щели в кутикуле, к перепончатому дну которых подходят чувствительные отростки нервных клеток, являются органами химического чувства и служат для обоняния. Органы зрения представлены простыми глазами, которые имеются у большинства паукообразных. Они расположены на дорзальной поверхности головогруди и обычно их бывает несколько: 12, 8, 6, реже 2. У скорпионов, например, имеется пара срединных более крупных глаз и 2-5 пар боковых. У пауков чаще всего 8 глаз, расположенных обычно в две дуги, причем средние глаза передней дуги крупнее остальных.
Скорпионы узнают себе подобных лишь на расстоянии в 2-3 см, а некоторые пауки - за 20-30 см. У пауков-скакунов (сем. Salticidae ) зрение играет особенно важную роль: если самцам замазать глаза непрозрачным асфальтовым лаком, то они перестают различать самок и производить характерный для периода спаривания "любовный танец".
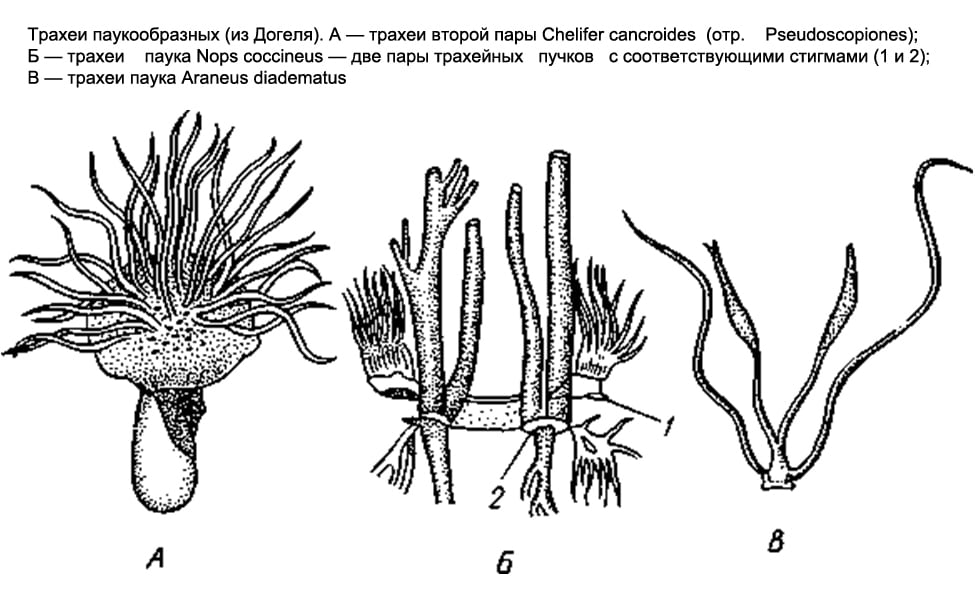
Органы дыхания Паукообразных разнообразны. У одних - это легочные мешки, у других - трахеи, у третьих и те и другие одновременно.
Только легочные мешки имеются у скорпионов, жгутоногих и у примитивных пауков. У скорпионов на брюшной поверхности 3-6-го сегментов переднебрюшия располагаются 4 пары узких щелей - дыхалец, которые ведут в легочные мешки. В полость мешка вдаются многочисленные параллельные друг другу листовидные складки, между которыми остаются узкие щелевидные пространства, в последние проникает через дыхательную щель воздух, а в легочных листочках циркулирует гемолимфа. У жгутоногих и низших пауков имеются лишь две пары легочных мешков.
У большинства других паукообразных (сольпуги, сенокосцы, лжескорпионы, часть клещей) органы дыхания представлены трахеями. На 1-2-м сегментах брюшка (у сольпуг на 1-м членике груди) имеются парные дыхательные отверстия, или стигмы. От каждой стигмы внутрь тела отходит пучок длинных, тонких, слепо замкнутых на концах воздухоносных трубочек эктодермального происхождения (образуются как глубокие впячивания наружного эпителия). У лжескорпионов и клещей эти трубочки, или трахеи, устроены просто и не ветвятся, у сенокосцев они образуют побочные веточки.
Наконец, в отряде пауков оба рода дыхательных органов встречаются совместно. У низших пауков, как уже отмечалось, имеются только легкие; в числе 2 пар они расположены на нижней стороне брюшка. У остальных пауков сохраняется лишь одна передняя пара легких, а позади последних имеется пара трахейных пучков, открывающихся наружу двумя стигмами. Наконец, у одного семейства пауков (Caponiidae ) легких нет вовсе, а единственные органы дыхания - 2 пары трахей.
Легкие и трахеи паукообразных возникли независимо друг от друга. Легочные мешки, несомненно, более древние органы. Считается, что развитие легких в процессе эволюции было связано с видоизменением брюшных жаберных конечностей, которыми обладали водные предки паукообразных и которые были похожи на жаброносные брюшные ножки мечехвостов. Каждая такая конечность впятилась внутрь тела. При этом образовалась полость для легочных листочков. Боковые края ножки приросли к телу почти на всем протяжении, кроме того участка, где сохранилась дыхательная щель. Брюшная стенка легочного мешка соответствует, следовательно, самой бывшей конечности, передний участок этой стенки - основанию ноги, а легочные листочки произошли от жаберных пластинок, находящихся на задней стороне брюшных ножек предков. Такое толкование подтверждается развитием легочных мешков. Первые складчатые зачатки легочных пластинок возникают на задней стенке соответствующих зачаточных ножек прежде, чем конечность углубится и превратится в нижнюю стенку легкого.
Трахеи возникли независимо от них и позже их как органы, более приспособленные к воздушному дыханию.
У некоторых мелких паукообразных, в том числе у части клещей, органы дыхания отсутствуют, и дыхание совершается через тонкие покровы.
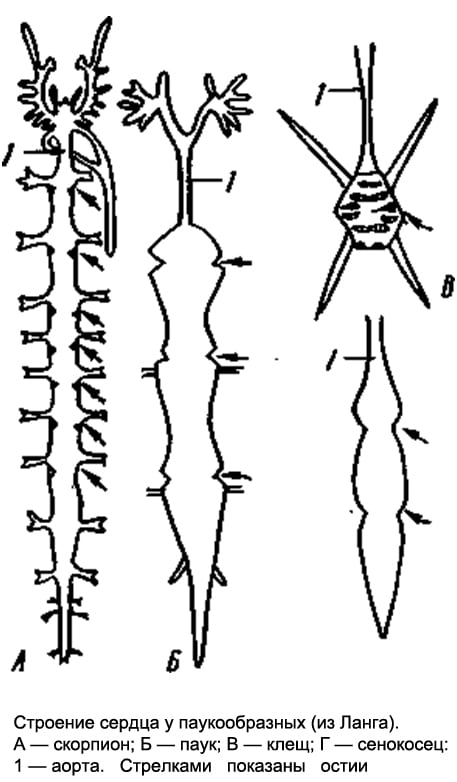
Кровеносная система. У форм с ясно выраженной метамерией (скорпионы) сердце представляет длинную трубку, залегающую в переднебрюшии над кишечником и снабженную по бокам 7 парами щелевидных остий. У других паукообразных строение сердца более или менее упрощается: так, у пауков оно несколько укорочено и несет всего 3-4 пары остий, а у сенокосцев число последних сводится до 2-1 пары. Наконец, у клещей сердце в лучшем случае превращается в короткий мешочек с одной парой остий. У большинства клещей в связи с их малыми размерами сердце совершенно исчезает.
От переднего и заднего концов сердца (скорпионы) или только от переднего (пауки) отходит по сосуду - передняя и задняя аорты. Кроме того, у ряда форм от каждой камеры сердца отходит по паре боковых артерий. Концевые веточки артерий изливают гемолимфу в систему лакун, т. е. в промежутки между внутренними органами, откуда она поступает в перикардиальныи участок полости тела, а затем через остии - в сердце. Гемолимфа паукообразных содержит дыхательный пигмент - гемоцианин.
Половая система. Паукообразные раздельнополы. Половые железы лежат в брюшке и в наиболее примитивных случаях парные. Очень часто, однако, происходит частичное слияние правой и левой гонад. Иногда у одного пола гонады еще парны, тогда как у другого слияние уже произошло. Так, у самцов скорпионов имеются два семенника (каждый из двух трубок, связанных перемычками), а у самок - один цельный яичник, состоящий из трех продольных трубок, соединенных поперечными спайками. У пауков в одних случаях гонады сохраняют обособленность у обоих полов, в других же у самки задние концы яичников срастаются, и получается цельная гонада. От гонад отходят всегда парные половые протоки, которые у переднего конца брюшка сливаются вместе и открываются наружу половым отверстием, последнее у всех паукообразных лежит на первом сегменте брюшка. У самцов имеются различные дополнительные железы, у самок нередко развиваются семяприемники.
Развитие. Вместо наружного оплодотворения, которые было свойственно далеким водным предкам паукообразных, у них развилось внутреннее оплодотворение, сопровождаемое в примитивных случаях сперматофорным осеменением или у более развитых форм копуляцией. Сперматофор представляет собой мешочек, выделяемый самцом, в котором находится порция семенной жидкости, защищенной таким образом от высыхания во время пребывания на воздухе. Самец у лжескорпионов и у многих клещей оставляет сперматофор на почве, а самка захватывает его наружными половыми органами. Обе особи при этом совершают "брачный танец", состоящий из характерных поз и движений. Самцы многих паукообразных переносят сперматофор в половое отверстие самки с помощью хелицер. Наконец, у некоторых форм имеются копулятивные органы, а сперматофоры отсутствуют. В ряде случаев для копуляции служат части тела, не связанные непосредственно с половой системой, например видоизмененные концевые членики педипальп у самцов пауков.
Большинство паукообразных откладывает яйца. Однако у многих скорпионов, лжескорпионов и у некоторых клещей наблюдается живорождение. Яйца большей частью крупные, богатые желтком.
У паукообразных встречаются различные типы дробления, однако в большинстве случаев имеет место поверхностное дробление. Позднее за счет дифференциации бластодермы образуется зародышевая полоска. Поверхностный слой ее образован эктодермой, более глубокие слои представляют мезодерму, а самый глубокий слой, прилегающий к желтку,- энтодерма. Остальная часть зародыша одета лишь эктодермой. Формирование тела зародыша происходит главным образом за счет зародышевой полоски.
В дальнейшем развитии следует отметить, что у зародышей сегментация бывает выражена лучше, а тело состоит из большего числа сегментов, чем у взрослых животных. Так, у зародышей пауков брюшко состоит из 12 сегментов, сходно со взрослыми ракоскорпионами и скорпионами, причем на 4-5 передних имеются зачатки ножек. При дальнейшем развитии все брюшные сегменты сливаются, образуя цельное брюшко. У скорпионов конечности закладываются на 6 сегментах переднебрюшия. Передняя пара их дает половые крышечки, вторая - гребенчатые органы, а развитие прочих пар связано с образованием легких. Все это указывает на то, что класс Arachnida произошел от предков с богатой сегментацией и с конечностями, развитыми не только на головогруди, но и на брюшке (переднебрюшии). Почти у всех паукообразных развитие прямое, но у клещей имеется метаморфоз.
Литература: А. Догель. Зоология беспозвоночных. Издание 7, переработанное и дополненное. Москва "Высшая школа", 1981